{kind=link}
Птахи́ (Aves) — клас теплокровних яйцекладних вищих хребетних тварин, які пересуваються на двох ногах, а їхні передні кінцівки перетворилися на крила. Станом на 2007 рік нараховують від 9.800 до 10.500 видів птахів. Вони населяють усі екосистеми Земної кулі від Арктики до Антарктики. Розміри видів коливаються від 5 см (колібрі) до 2,75 м (страус). Птахи — це спеціалізована гілка хребетних, що виникла від тероподних динозаврів у юрському періоді (близько 160 млн років тому).
Зміст
2. Систематика
3. Поширення
4. Особливості будови
- Загальна характеристика птахів
- Оперення та роговий покрив
- Розміри
- Скелетна система
- Склад скелету
- Луски
- Дихальна система
- Кровоносна система
- Травна система
- Органи виділення
- Репродуктивна система
- Нервова система
- Органи чуття
6. Голосовий апарат та його роль
- Вокалізація птахів
- Визначення
- Анатомія
- Соціальні аспекти та етологія
- Функція
- Мова та характеристики вокалізації
- Діалекти
- Навчання та контроль
- Загальні характеристики
- Привласнення та імітація
- Нейрофізіологія
- Ідентифікація і систематика
- Спів птахів і музика
7. Пересування
- Історія дослідження
- Еволюція
- З дерев униз
- Із землі вгору
- Біг за допомогою крил
- Втрата польоту у деяких сучасних птахів
- Нелітаючі птахи
- Анатомія структур, пристосованих до польоту
- Скелет крила
- Пір'я крил
- Махові пера
- Махові пера крил
- Першорядні махові
- Другорядні махові пера
- Третьорядні махові пера
- Виїмки
- Придаткове крило
- Затримка розвитку в гоацинів
- Крильце
- Типові форми крил
- Хвіст
- Інші адаптації до польоту
- Режими польоту
- Зліт
- Посадка
- Маховий політ
- Переривистий політ
- Політ упорядкованою групою
- Зависання
- Зависання проти вітру
- Ширяння
- Термальне ширяння
- Маневрування
8. Поведінка
9. Міграції
- Типи переміщень птахів
- Осілі птахи
- Міграція на великі відстані
- Кочівля та міграція на короткі відстані
- Вторгнення і розосередження
- Фізіологія і контроль
- Час міграції
- Орієнтація і навігація
- Помилки при міграції
- Штучне управління міграцією
- Еволюція та екологія виникнення міграції
- Екологічні наслідки перельотів птахів
- Методи дослідження
- Дослідження орієнтації
- Загрози та збереження птахів
- Міжнародний день мігруючих птахів
10. Живлення
11. Значення в природі та житті людини
- Взаємовідносини з людиною
- Економічне значення
- Роль у природних екосистемах
- Релігія, фольклор і культура
12. Охорона птахів
Характерними рисами сучасних птахів є наявність пір'я, дзьоба, позбавленого зубів, відкладання яєць з міцними оболонками, високий рівень метаболізму, чотирикамерне серце, легкий, але міцний скелет. Усі сучасні птахи мають крила, що розвинулися з передніх кінцівок. Винятком є нещодавно вимерлий птах моа з Нової Зеландії. Більшість птахів здатні до польоту, до нелітаючих належать страусоподібні, пінгвіни та невелика кількість ендемічних острівних видів. Птахи також мають пристосовану до польоту травну і дихальну системи. Деякі види, особливо воронові та папуги, належать до тварин із найрозвиненішим інтелектом, здатних застосовувати знаряддя для добування їжі. Багато птахів уміють передавати індивідуальний досвід між поколіннями.
Значна частина птахів щороку здійснюють дальні міграції, а багато нерегулярно кочують на короткі відстані. Птахи є соціальними тваринами, які застосовують для комунікації візуальні і звукові сигнали, зокрема спів; для них характерна соціальна поведінка, що включає утворення зграй, спільне вигодовування пташенят, полювання та захист від ворогів. Переважна більшість птахів є моногамними — вони утворюють пари, зазвичай, на один сезон гніздування, інколи на декілька років або на все життя. В інших видів спостерігається полігінія («багато самиць») або, рідко, поліандрія («багато самців»). Яйця здебільшого відкладаються в особливим чином зроблені гнізда та висиджуються партнерами. Багато птахів досить довгий період після вилуплення з яєць пташенят піклуються про них.
Багато видів мають важливе економічне значення для людини, здебільшого як джерело їжі, отриманої в результаті ведення сільського господарства або полювання. Деякі види, особливо співочі птахи і папуги, є популярними хатніми тваринами. Птахи посідають чільне місце в усіх сферах людської культури — від релігії до поезії та музики. Внаслідок антропогенного впливу за період із XVII ст. вимерло близько 120–130 видів птахів. На початку XXI ст. близько 1200 видів перебувають під загрозою зникнення через діяльність людини, попри зусилля щодо їх збереження.
В Україні налічують 423 або 428 видів. Серед них 267–270 гніздові, з яких 132–138 зимуючі; 17 з'являються лише на зимівлі, 129 спостерігаються лише у період сезонних міграцій або мають статус залітних.
Науку, що вивчає птахів, називають орнітологією.
Походження і еволюція
Загальновизнано, що птахи походять від Архозаврів — підкласу плазунів, що панував у мезозойську еру. Теорія еволюції птахів від плазунів виникла після відкриття 1860 року скам'янілих решток археоптерикса — тварини, що існувала близько 150 млн років тому у верхній юрі. Археоптерикс мав одночасно ознаки як плазунів, так і птахів. Спільним з плазунами була особлива будова таза і ребер, зуби, лапи з кігтями та довгий хвостовий відділ хребта. У той же час знайдені скам'янілі рештки продемонстрували добре збережені відбитки крил із пір'ям, подібним до сучасного пташиного. В останнє десятиріччя більшість дослідників не розглядають археоптерикса як спільного предка усіх сучасних птахів, однак вважають його примітивним найдавнішим представником класу Aves, який, імовірно, близько споріднений зі справжнім предком.
{kind=link}
Археоптерикс — один із можливих предків птахів
На сьогодні найпоширенішою є гіпотеза походження птахів від тероподних динозаврів.
Починаючи з 1980-х років кількість знайдених решток мезозойських птахів значно зросла, проте точних предків цих тварин не встановлено. Ситуація ускладнюється тим, що важко визначити ознаки, за якими птахів можна відрізнити від тероподних динозаврів. Якщо раніше однією з найважливіших ознак була наявність пір'яного покриву, то низка знахідок наприкінці ХХ — на початку XXI ст. у провінції Ляонін на північному сході Китаю свідчать, що дрібні тероподи мали пір'я, не пристосоване до польоту.
Одним із скептиків тероподової теорії був російський вчений Є. М. Курочкін. Він вважав, що тільки викопні Enantiornithes та археоптерикс походять від тероподів, а сучасні птахи — від якихось інших архозавроподібних, подібних до протоавіса.
Нові представники віялохвостих птахів відомі з крейдового періоду. Першою широко поширеною групою птахів стали енанціорніси, які захопили різноманітні екологічні ніші — від мешканців піщаного морського узбережжя й мисливців за рибою до форм, що мешкали на деревах та живилися насінням. Більш прогресивно розвиненими були іхтіорніси, які за зовнішнім виглядом були схожі на мартинів, мешкали на морських просторах та живилися рибою. Морські птахи з ряду гесперорнісоподібних із мезозойської ери були добре пристосовані до полювання на рибу, вони втратили здатність до польоту та повністю перейшли до водного способу життя. У ранньокрейдовому періоді (близько 120 млн років тому) існували також конфуціорніси (Confuciusornis), рештки яких знайдено в Китаї. Це були перші птахи, що мали дзьоб, крім того, вони були беззубими, на відміну від гесперорнісів та іхтіорністів.
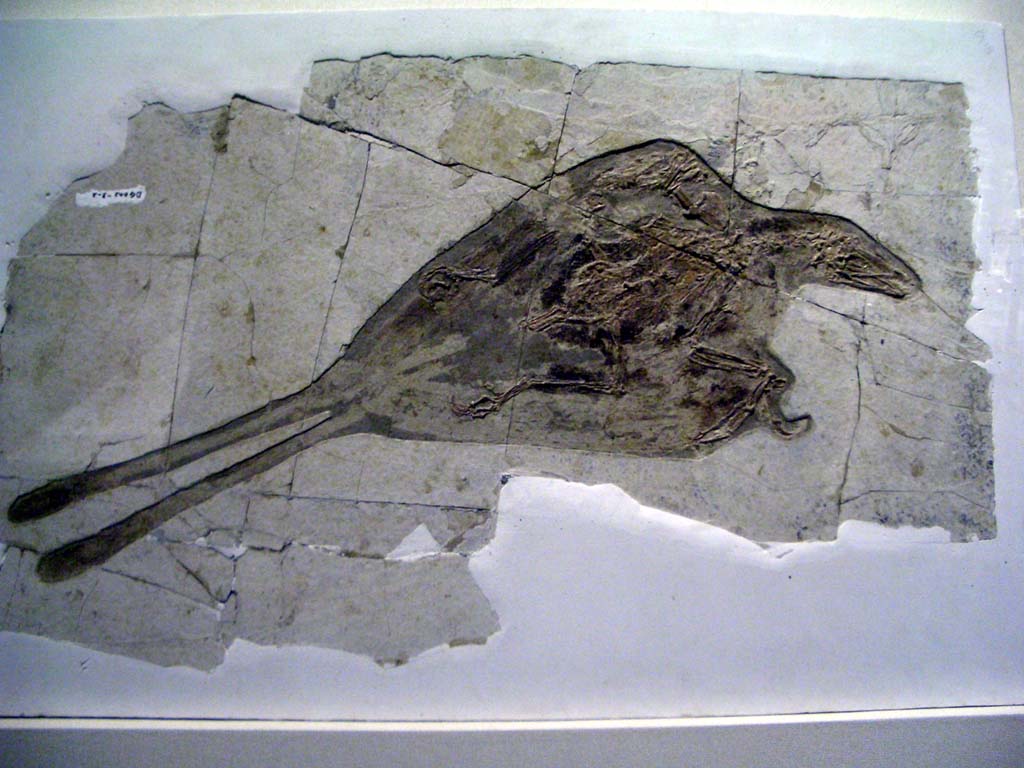{kind=link}
Конфуціусорніси — перші птахи, що були беззубими та мали дзьоб
Першу спробу побудови системи тварин здійснив у 4 ст. до н. е. грецький вчений Аристотель. У його працях згадується близько 160 видів птахів. Початок наукової систематики було закладено відомою працею шведського натураліста Карла Ліннея «Система природи» (1735–1758). Він виділив 6 класів, 65 родів та 554 види птахів. Були введені ієрархічні таксономічні категорії та бінарна номенклатуру для позначення видів, які застосовують і досі. У системі Ліннея птахам був наданий ранг класу.
Основу сучасних уявлень про систему класу було закладено класичними роботами Макса Фюрбрінгера (Furbringer, 1888) та Ганса Гадова (Gadow, 1891–1893). На відміну від своїх попередників, обидва автори для аналізу спорідненості видів використали великий за обсягом порівняльно-анатомічний матеріал. У подальшому багато авторів, по суті удосконалювали цю систему птахів.
Одна з сучасних альтернативних філогенетичних систем розглядає групу «Aves» як кладу тероподних динозаврів (Livezey, Zusi, 2007).
Деякі відомі фахівці з динозаврів, такі як Жак Готьє, Луїс Кіаппі, під категорією «Aves» розуміють лише сучасних птахів, не включаючи туди групи, відомі лише за викопними рештками — археоптерикса, конфуціорніса, патагопрерискса та деяких інших. Однак ці вчені ввели нову категорію «Avialae», або «птахів у широкому розумінні», яка об'єднує як сучасні види, так і їх викопних попередників.
Усі сучасні птахи належать до інфракласу віялохвостих птахів (Neornithes), які протиставляються ящерохвостим (Archaeornithes). До останніх належить лише один вид — археоптерикс. Віялохвості, у свою чергу, розділяються на дві групи — Безкілеві (Paleognathae) та Кілегруді (Neognathae). Зазвичай ці групи мають ранг надряду, хоча в роботах Лайвезі та Зусі розглядаються як когорта.
Страус — найбільший серед сучасних птахів
Пінгвін Аделі — належить до Кілегрудих, проте не здатен до польоту
Ластівка берегова — один з видів-космополітів
Залежно від класифікації серед сучасних птахів налічують від 9 800 (Clements, 2007) до 10 500 (Frank, 2006) видів.
Нижче наведено традиційну класифікацію (так званий порядок Клементса), що включає 29 рядів:
Підклас Віялохвості птахи (Neornithes)
Надряд Безкілеві (Paleognathae)
Страусоподібні (Struthioniformes) — страуси, ківі, казуари, нанду
Тинамуподібні (Tinamiformes)
Надряд Кілегруді (Neognathae)
Гусеподібні (Anseriformes)
Куроподібні (Galliformes)
Сивкоподібні (Charadriiformes) — мартини, кулики, алькові
Гагароподібні (Gaviiformes)
Пірникозоподібні (Podicipediformes)
Буревісникоподібні (Procellariiformes) — альбатроси, буревісники
Пінгвіноподібні (Sphenisciformes)
Пеліканоподібні (Pelecaniformes) — пелікани, баклани, олуші, фрегати
Фаетоноподібні (Phaethontiformes)
Лелекоподібні (Ciconiiformes) — лелеки, чаплі та інші
Американські грифи (Cathartiformes)
Фламінгоподібні (Phoeriicopteri)
Соколоподібні (Falconiformes) — яструби, соколи та інші
Журавлеподібні (Gruiformes) — журавлі, дрохви, пастушки та інші
Рябкоподібні (Pteroclidiformes)
Голубоподібні (Columbiformes)
Папугоподібні (Psittaciformes)
Зозулеподібні (Cuculiformes) — зозулі, турако
Гоациноподібні (Opisthocomiformes)
Совоподібні (Strigiformes)
Дрімлюгоподібні (Caprimulgiformes)
Серпокрильцеподібні (Apodiformes) — серпокрильці, колібрі
Сиворакшеподібні (Coraciiformes)
Дятлоподібні (Piciformes)
Трогоноподібні (Trogoniformes)
Птахи-миші (Coliiformes)
Горобцеподібні (Passeriformes) — найрізноманітніший ряд
{kind=link}
Страус — найбільший серед сучасних птахів
{kind=link}
Пінгвін Аделі — належить до Кілегрудих, проте не здатен до польоту
Птахи поширені на Земній кулі практично повсюдно. На Крайній Півночі між 80 та 83° пн. ш. місцями гніздяться біла і тундряна (Lagopus mutus) куріпки, крем'яшник, плоскодзьобий плавунець, полярний крячок, морянка, пуночка та інші птахи. Зальотних пуночок, чистиків, білих мартинів (Pagophila eburnea) та полярних мартинів (Larus glaucoides) спостерігали на 88—89° пн. ш.
Птахи трапляються повсюдно на материках, включаючи найбільші безводні пустелі й гори до смуги вічних снігів: на висоті 5500—6500 м гніздяться ягнятники, грифи, крук, клушиця (Pyrrhocorax pyrrhocorax), біла плиска, гімалайська тинівка (Prunella himalayana) та інші, а під час міграцій зграї пролітаючих гусей, журавлів та навіть горобцеподібних іноді помічали на висотах 7000—8000 м н.р.м. Заселяють птахи також найвіддаленіші від материків океанічні острови.
На краях Антарктичного щита місцями гніздяться пінгвіни (імператорський, Аделі, антарктичний), сніжний і гігантський буревісники (заліт останнього помічено за 80 км від Південного полюса) та інші.
Лише поодинокі види є космополітами — широко поширені на всіх материках (крім Антарктиди). Це сапсан та скопа. Дещо менший ареал у сипухи, морського зуйка, сільської ластівки та деяких інших птахів. Поширені деякі синантропні види.
Існують види з дуже невеликими ареалами. Деякі види колібрі мешкають лише на окремих горах в Андах. Територією окремих невеликих островів обмежене поширення частини галапагоських в'юрків, гавайських квіткарок (Drepanididae) та деяких інших птахів.
Ареали окремих видів суттєво змінюються під впливом різноманітних причин.
Найрізноманітнішою фауна птахів є в тропіках — тут зустрічається близько 85% сучасних видів та підвидів, тоді як у помірних та холодних широтах — лише 15%. Більша, ніж на інших континентах, кількість ендемічних видів спостерігається в Південній Америці.
Загальна характеристика птахів
{kind=link}
Зовнішня будова типового птаха: 1 — дзьоб, 2 — голова, 3 — райдужка, 4 — зіниця, 5 — мантія, 6 — малі криючі крила, 7 — крило, 8 — криючі другорядних махових, 9 — криючі першорядних махових, 10 — надхвістя, 11 — першорядні махові, 12 — підхвістя, 13 — стегно, 14 — гомілково-тарзальні зчленування, 15 — цівка, 16 — пальці, 17 — гомілка, 18 — черево, 19 — боки, 20 — груди, 21 — горло, 22 — «вуса»
Однією з головних особливостей птахів порівняно з іншими групами хребетних є здатність до польоту. Існує лише невелика кількість (близько 60 видів) нелітаючих або майже нелітаючих птахів, однак усі вони в процесі еволюції так чи інакше втратили цю функцію, наявну в їхніх предків. Вміння пересуватися в повітрі визначає усю біологію цього класу тварин, у тому числі — будову організму.
Тіло птахів компактне, обтічної форми. Шия в більшості видів тонка та гнучка, що забезпечує рухливість голови. На голові розташовано дзьоб, утворений наддзьобком та піддзьобком. Рух у повітрі здійснюється за допомогою передніх кінцівок, які перетворилися на крила, а також хвоста. Крила слугують як аеродинамічними поверхнями, що утримують птаха в повітрі, так і джерелом тягової сили для руху вперед. Скелет птахів легкий і міцний, завдяки пневматизації кісток та їхнього зростання.
Ноги птахів мають 4 відділи: стегно, гомілку, цівку та пальці, яких зазвичай чотири, хоча інколи буває три або навіть два (африканський страус). У більшості випадків три з чотирьох пальців обернені вперед і один — назад. Ноги птахів приймають усю вагу тіла під час пересуванні на землі, на зльоті та під час посадки, лазіння по деревах. Під час пересування на землі стегнові кістки малорухливі і залишаються в майже горизонтальному положенні. Саме фіксація стегнової кістки дає змогу підтримувати черевний повітряний мішок під час вдиху, що визначає роботу дихальної системи птахів та дає змогу їм літати. Це одна з головних характеристик птахів, усі інші тварини, що пересуваються на землі, мають рухливий тазостегновий суглоб. Іншою особливістю ніг птахів є добрий розвиток м'язів, що не характерно для інших літаючих тварин.
Іншими важливими пристосуваннями до польоту є виникнення оперення, чотирикамерного серця, подвійного дихання, висока стала температура тіла, високий рівень метаболізму, зникнення зубів та сечового міхура, розмноження шляхом відкладання яєць.
{kind=link}
Будова пташиного пера (на прикладі махового): 1 — опахало, 2 — стовбур, 3 — борідки, 4 — пухова частина, 5 — очин
Перо́ — рогове утворення шкіри птахів. Пера вкривають більшу частину поверхні тіла птаха та формують оперення.
Структура
Найскладніша будова притаманна контурним перам, що утворені стрижнем та двома опахалами, розташованими в одній площині. Нижня частина стрижня — так званий очин — порожня та позбавлена опахал, всередині розташовано плівчасте утворення, що називається «душка». Решта стрижня — «ствол» — має серцевину, що складається з легкої рогової комірчастої тканини.
Опахала утворені довгими борідками, від яких відходять менші борідки другого порядку; на них, в свою чергу, розташовані рогові гачки, якими зкріплюються борідки пера. В разі відсутності гачків борідки залишаються незчіпленими, і такий вид пера називається пуховим.
Завдяки оперенню птахи можуть набувати різного забарвлення, що досягається двома шляхами. По-перше, шляхом вміщення в борідках пера барвників різного кольору (наприклад, у папуг); і по-друге, завдяки специфічній структурі поверхні пера, що забезпечує відбивання світла разом з його спектральним розкладенням (наприклад, у павича). Птахи, яким притаманне таке «структурне» забарвлення, втрачають його в разі намокання пір'я.
Різновиди
Розрізняють такі види пер: контурні, нитчасті, пухові, пух та щетинки.
До контурних пер належать: махові, що відіграють головну роль в утворенні несучої поверхні крила; рульові, що утворюють хвіст, та криючі, що вкривають тулуб птаха і значну частину крил.
У більшості птахів (за деякими винятками, наприклад, пінгвінів) криючі пера вкривають тіло не скрізь: ділянки шкіри, де ростуть пера (птерилії) чергуються з ділянками, де пера відсутні (аптерії). Аптерії вкриті перами з сусідніх птерилій.
Нитчасті пера відрізняються довгим, тонким та м'яким стволом, та дуже нечисленним борідками другого порядку на його дистальному кінці. Звичайно, вони вкриті контурними перами, але у деяких птахів (наприклад, у бакланів) на ділянках шиї та потилиці виступають на поверхню оперення.
Пухові пера, як і пух, характеризуються м'яким та тонким стрижнем і незчепленими борідками. Ці види пер запобігають охолодженню тіла.
Для пуху характерний вкорочений стрижень та незчеплені борідки першого порядку, за майже повної відсутності борідок другого порядку. Ембріональне перо, з якого розвиваються всі форми пер, за будовою подібне до пуху.
Щетинки являють собою стрижні пер, позбавлені борідок. Вважається, що вони, як і нитчасті пера, виконують функцію органів дотику.
Пера на кінцях з часом стираються, розтріпуються та вицвітають, що призводить птахів до необхідності линяння, тобто періодичної зміни пір'я.
У великих за розміром птахів загальна кількість пір'їн може досягати 25 тис. (лебідь). Відносна маса оперення становить 4,6—9,5% від маси тіла.
Призначення оперення різноманітне. Воно забезпечує можливість польоту, утворюючи несучі поверхні (крила, хвіст) та створюючи обтічність тіла. Виконує теплоізолюючу та водовіштовхуючу функції. Завдяки забарвленню оперення виникає характерний вигляд птаха, що полегшує розпізнавання особин свого виду. У частини видів забарвлення оперення допомагає маскуванню, роблячи птаха малопомітним на фоні субстрату.
Походження та розвиток
Тривалий час вважалося, що еволюційно пера походять від шкірних лусок рептилій, але останнім часом цей постулат поставлено під сумнів. Спочатку пера виконували функцію теплоізоляції та, вірогідно, маркерну, у шлюбній поведінці, і лише потім стали пристосуванням для польоту.
Експериментально доведено, що відсутність в організмі ембріону птахів білкового рецептора, яка спричинює утворення перетинок між пальцями ніг, викликає також і перетворення шкірної луски на ногах на пера.
Птахи щоденно ретельно доглядають за оперенням, на що витрачають у середньому 9% свого денного часу. Догляд за оперенням у птахів полягає у змащуванні його секретом куприкової залози або так званою «пудрою», що утворюється зі спеціального пуху. Зміна оперення відбувається внаслідок линяння, у певні пори року. У дорослих птахів майже всіх видів протягом року буває одне повне линяння.
Механізм розвитку пера
В результаті розростання клітин епідермісу та кутису на шкірі утворюється горбок (він подібний до зачатку луски плазунів), що поступово розростається у вигляді спрямованого назад виросту, основа якого заглиблюється в шкіру, утворюючи піхву пера. Сполучнотканинна частина виросту перетворюється в пронизаний кровоносними судинами сосочок пера, що росте. Одночасно епідермальний шар, що розростається - диференціюється на тонкий роговий чохлик, який закриває перо, що росте, а з підлягаючих клітин, які посилено діляться та поступово роговіють, формується стержень з борідками, що відходять від нього. В міру росту пера оточуючий його роговий чохлик поступово злущується, а борідки розпрямляються, утворюючи опахало. Сосочок засихає; від нього в очині пера залишається лише плівчаста душка.
Використання людиною
Пера застосовують для набивки перин, подушок, м'яких меблів, а також для утеплення одягу (особливо цінується пух гаги).
У багатьох народів (зокрема, до середини 19-го сторіччя і в західноєвропейських країнах) пера входять до складу традиційного святкового вбрання.
Розміри птахів обмежуються здатністю до польоту, тому варіюють у невеликих межах. Маса найбільших літаючих птахів не перевищує 14—16 кг (лебеді, грифи, дрохви) при розмаху крил до 3—4 м (пелікани, альбатроси). Найдрібніші серед птахів — деякі колібрі з максимальною масою 1,6—2 г. Втрата здатності до польоту частково призводить до збільшення розмірів: маса великих пінгвінів досягає 40 кг, казуарів та африканських страусів — 80—100 кг. Деякі з вимерлих птахів досягали маси 300–400 кг (моа, епіорніси).
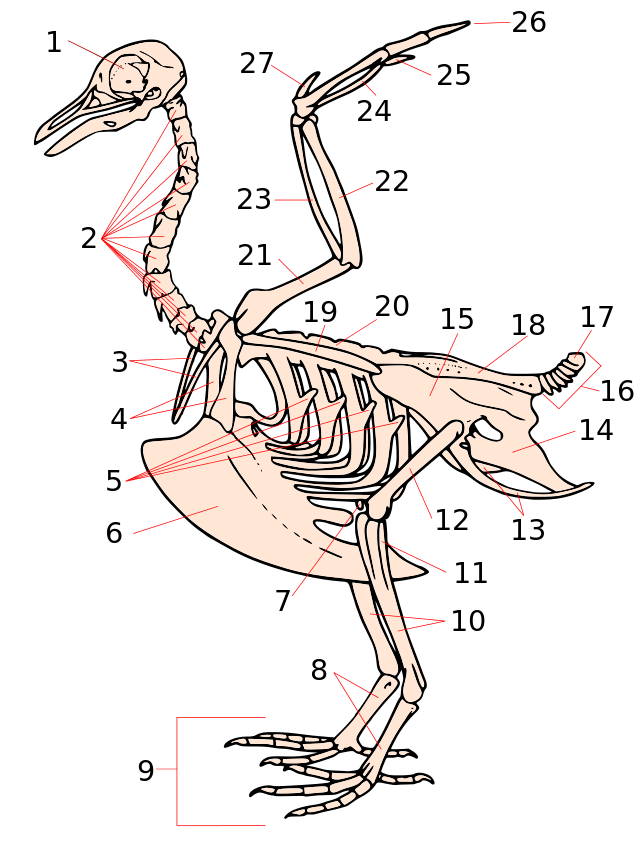{kind=link}
Скелет птаха на прикладі голуба. 1 — череп, 2 — шийні хребці, 3 — вилочка, 4 — коракоїд, 5 — гачкоподібні вирости ребер, 6 — кіль, 7 — колінна чашечка, 8 — цівка, 9 — пальці, 10 — велика гомілкова кістка (тибіотарсус), 11 — мала гомілкова кістка (тибіотарсус) 12 — стегно, 13 — сіднична кістка (тазова кістка), 14 — лобкова кістка (тазова кістка), 15 — клубова кістка (тазова кістка), 16 — хвостові хребці, 17 — пігостиль, 18 — складний криж, 19 — лопатка, 20 — поперекові хребці, 21 — плечова кістка, 22 — ліктьова кістка, 23 — променева кістка, 24 — п'ястно-зап'ястна кістка, 25—27 — пальці.
Основні властивості скелета птахів визначаються пристосуванням до польоту. Це виявляється перш за все в його легкості та міцності. Легкість скелета обумовлена тонкістю та пневматичністю кісток (включаючи кістки черепа). Внаслідок редукції кісткового мозку, більшість кісток трубчасті, заповнені повітрям, що потрапляє з повітряних мішків. Міцність скелета обумовлюється зростанням окремих кісток, їх структурою й наявністю в їх складі значної кількості неорганічних солей. Пристосування до польоту призвело також до зменшення загальної кількості кісток внаслідок злиття (на ранніх стадіях розвитку) або редукції окремих кісткових елементів. Відносна вага скелета птахів порівняно менша, ніж у ссавців і дорівнює 7,1—11,7% маси тіла. Легкість кісток дала змогу птахам різко видовжити передні та задні кінцівки (вони перевищують довжину тулуба в 2—3 рази, а у деяких — і більше), не збільшуючи відносної маси скелета.
Скелет птахів можна поділити на череп, хребет, кінцівки та їх пояси.
Череп птахів схожий на череп плазунів, але відрізняється легкістю; збільшенням об'єму черепної коробки, у зв'язку з сильним розвитком головного мозку; великими очними западинами; розташуванням потиличного отвору на дні черепа. У дорослих птахів усі кістки зливаються до повного зникнення швів і весь череп є суцільною тонкостінною, але міцною кістковою коробкою. Він складається з п'яти головних кісток: лобної (верхня частина голови), тім'яної (задня частина голови), передщелепної та носової (безпосередньо над дзьобом) і нижньощелепної кістки (безпосередньо під дзьобом). Череп сучасних птахів позбавлений зубів, їхню функцію частково виконує дзьоб. Череп більшості птахів важить близько 1% від повної маси тіла.
Кількість хребців у різних птахів коливається в межах 39—63. Хребці є гетероцельними (тобто мають сідлоподібні поверхні сполучення).
Хребет птахів складається з 5 відділів: шийного, грудного, поперекового, крижового і хвостового.
Шийний відділ хребта птахів містить 13—25 хребців, що більше ніж у будь-яких інших тварин, внаслідок чого в більшості з них дуже гнучка шия.
Грудний відділ складається з 3—10 хребців, які зростаються між собою, утворюючи суцільну кістку. До кожного грудного хребця приєднуються пара ребер, які доходять до грудини і рухомо з'єднуються з нею. Кожне ребро складається з рухомо з'єднаних кісткових відділів: спинного і черевного. Спинний відділ має гачкоподібний відросток, який вільно налягає на сусіднє заднє ребро. Грудна кістка у птахів досить масивна, на її середній частині у всіх птахів, крім бігаючих, розташовано великий гребінь, або кіль, до якого прикріплюються міцні м'язи, що рухають крила.
Поперекові, крижові та частина хвостових хребців утворюють характерний для птахів складний криж (лат. synsacrum ). Разом з тазовим відділом він надає міцної опори заднім кінцівкам, що вкрай важливо у зв'язку з перенесенням ваги тіла на них під час руху суходолом.
Хвостовий відділ має 6—11 вільних хребців та закінчується куприковою кісткою (пігостиль), яка утворилася внаслідок злиття кількох хвостових хребців.
Передні кінцівки перетворюються на крила. Плечова кістка сполучається з променевою й ліктьовою кістками, що формують передпліччя. Зап'ястя і п'ястя утворені п'ястно-зап'ястною кісткою. Скелет пальців редукується: добре розвинені тільки дві фаланги ІІ пальця та по одній фаланзі від І та ІІІ пальців. Кістки крила дуже легкі, що полегшує політ.
Пояс передніх кінцівок складаються зі зрослих ключиць (так звана вилочка), парних коракоїдів та лопаток.
До складу поясу задніх кінцівок входять тазові кістки: клубова, сіднична і лобкова, злиті в одну безіменну кістку.
Нижня кінцівка складається із стегнової кістки, яка у колінному суглобі сполучається з тибіотарсусом (гомілкою) і малою гомілковою кісткою (на бічній стороні ноги). Злиті плесно і передплесно (плеснопередслесно) формують верхню частину ступні — цівку, до якої прикріплені кістки пальців. Кістки ніг птахів найважчі, що приводить до низького центра маси тіла та допомагає у польоті. Проте загалом скелет становить лише 5% від повної маси тіла.
Луски птахів складаються з того ж самого позаклітинного кератину, як і дзьоби, кігті та шпори. Вони знайдені головним чином на пальцях та біля основи ніг, але інколи можуть знаходитися і вище, до щиколотки у деяких птахів. Луски більшості птахів практично не перекриваються, за виключенням лусок зимородків і дятлів. Вважається, що луски птахів гомологічні лускам плазунів та ссавців.
Ембріони птахів починають розвиток з утворення гладкої шкіри. Проте пізніше, зовнішній шар шкіри ніг, роговий шар, може кератинизуватися, потовщитися та утворити луски. Ці луски можуть бути організовані у кілька типів структур:
- Cancella — маленькі луски, що представляють собою лише незначне потовщення шкіри та утворення жолобків на її поверхні.
- Reticula — маленькі, але чіткі та окремі луски. Знайдені на внутрішньому за зовнішньому боках плесна.
- Scutella — луски середнього розміру, знайдені на задній частині плесна.
- Scute — найбільші за розміром луски, зазвичай на передній стороні плесна та задній стороні пальців.
На ногах деяких птахів луски перемежаються пір’ям. Цибулини пір'я можуть знаходитися між лусками або безпосередньо під ними, в глибших шарах шкіри. В цьому разі пір'я може виходити назовні крізь луски.
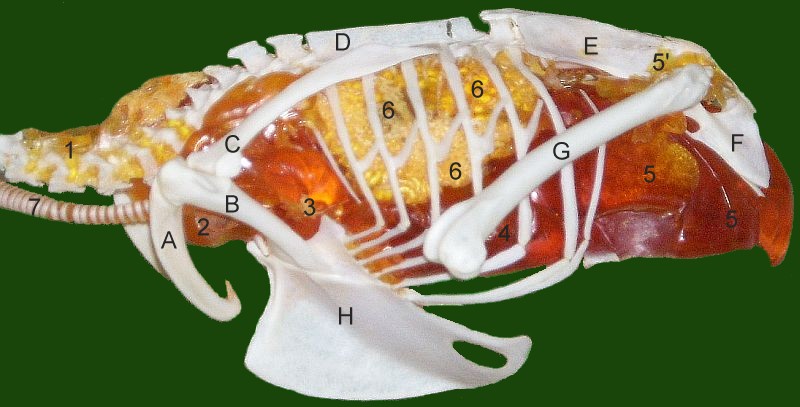{kind=link}
Система циркуляції повітря звичайного боривітра. Потік повітря завжди відбувається зліва (з постеріорного кінця) направо через легені як протягом вдожу, так і протягом видоху. 1 шийний повітряний мішок, 2 ключичний повітряний мішок, 3 черепний thoracal , 4 хвостово-грудний повітряний мішок, 5 черевний повітряний мішок, 6 легені, 7 трахея.
Через високий рівень метаболізму, потрібний для польоту, птахи вимагають високого рівня постачання кисню. Через це, розвиток ефективної дихальної системи відбувся одночасно з еволюцією польоту в птахів. Птахи забезпечують постачання повітря до легенів за допомогою повітряних мішків, унікальних для птахів та, можливо, динозаврів. Ці мішки не грають прямої ролі в газообміні, проте зберігають повітря і діють подібно до міхів, дозволяючи легеням отримувати постійний потік повітря.
У птахів за дихання відповідають три окремих групи повітряних мішків: передні повітряні мішки (ключичний, шийні і передні грудні), легені і задні повітряні мішки (задні грудні та черевні). Передні та задні повітряні мішки (всього зазвичай дев'ять) розширяються під час вдоху. Повітря потрапляє до тіла птаха через трахею, половина потрапляє до задніх повітряних мішків, а половина проходить через легені та потрапляє до передніх повітряних мішків. Під час видиху мішки стискаються, при цьому повітря з передніх повітряних мішків виходить прямо через трахею, а повітря із задніх спочатку проходить через легені.
Через те, що свіже повітря проходить через легені завжди в одному напрямку, у них не відбувається змішування багатого та бідного на кисень (і багатого на вуглекислий газ) повітря, як це відбувається в легенях ссавців. В результаті парціальний тиск кисню в легенях птахів такий же як і в навколишньому середовищі, що дозволяє ефективніше постачати кисень, ніж це відбувається у ссавців.
На відміну від легенів ссавців, легені птахів не мають альвеол, проте містять мільйони тонких проток — парабронхів, що сполучаються з дорсобронхами та вентобронхами. Повітря потрапляє через стінки парабронхів до газових везикул, атріїв, через радіальні виступи на поверхні парабронхів. Отриманий кисень потрапляє до капілярів, де за допомогою дифузії і відбувається газообмін.
Травна система починається з ротового отвору. Велике значення для захоплення їжі відіграє дзьоб, форма та розміри якого значно варіюють у різних видів, що залежить від способу живлення. Ротова порожнина переходить у стравохід. Останній у багатьох видів має розширення — воло, в якому їжа накопичується. Розмір та форма вола досить сильно відрізняються між видами птахів. Члени ряду голубоподібних, такі як голуби, здатні виділяти їстивне голубине або волове молоко, яким вони годують пташенят. Характерною особливістю травної системи є наявність складного шлунка з двох відділів — залозистого і м'язового. Другий шлунок деяких видів містить невеликі шматочки гравію та дрібного каміння, яке ці птахи заковтують для допомоги травленню, і яке виконує функцію зубів ссавців та плазунів. Використання другого шлунка однаково у птахів та динозаврів, в скам'янілостях яких залишилося каміння, так званий гастороліт. Кишечник поділяється на тонкий та товстий. Розвинені травні залози — печінка та підшлункова, протоки яких відкриваються в дванадцятипалу кишку. Інтенсивність травлення у птахів дуже висока. Наприклад, у горобців, від заковтування гусені до виведення її неперетравлених решток з екскрементами минає лише 15—20 хвилин, жуків — близько 1 години та насіння — 3—4 години.
{kind=link}
У омелюха з'їдені ягоди горобини проходять весь кишечник за 8—10 хв.
Кровоносна система має два кола кровообігу, артеріальна і венозна кров не змішується, ця адаптація дозволяє ефективне постачання поживних речовин та кисню до всіх тканин тіла, забезпечуючи високий рівень активності. Серце чотирикамерне, з однією правою дугою аорти. Відрізняється високою частотою серцевого ритму, особливо у дрібних птахів, — так серце червоногорлого колібрі б'ється з частотою 1200 ударів за хвилину (або 20 ударів за секунду).
Органи виділення — тазові нирки (метанефрос), від яких відходять сечоводи, що відкриваються в клоаку. Основним продуктом азотистого обміну птахів є сечова кислота, у зв'язку з чим сечовий міхур редукується.
Репродуктивна система. Хоча більшість птахів не мають зовнішніх статевих органів, самець має два сім'яники, що збільшуються у сотні разів під час шлюбного сезону, коли вони починають виробляти сперму. Яєчники самок також збільшуються у розмірі, хоча зазвичай лише лівий яєчник має повну функціональність. Але якщо лівий яєчник пошкоджується з результаті хвороби або інших проблем, правий яєчник може взяти на себе його функцію. Якщо ж він не здатний відновити функцію, самки деяких видів птахів можуть розвинути вторинні статеві ознаки самців, а інколи навіть зміну у голосі.
Більшість видів птахів на мають статевого члена, у них сперма до спаровування зберігається в сім'яних клубочках в межах опуклості клоаки. Під час спаровування самка відхиляє свій хвіст убік, а самець сідає на самку зверху, розташовується спереду (у Notiomystis cincta) або іншим чином присувається дуже близько до неї. Клоаки птахів торкаються таким чином, що сперма може потрапити до статевого тракту самки. Зазвичай це трапляєтсья швидко, часто за менш ніж пів секунди.
У тілі самки сперма зберігається у призначених для цього трубках, де вона може знаходитися від тижня до року, залежно від виду. Кожне яйце запліднюється окремо, коли воно залишає яєчник, але перед відкладанням. Після відкладення яйце продовжує розвиток вже за межами тіла самки.
Багато водоплавних птахів та деякі інші види, такі як страус та індичка, мають статевий член. Поза часом спаровування він схований у проктодеумі, відділі клоаки.
Після насиджування яєць та вилуплення пташенят, батьки забезпечують їх різним ступенем догляду та захисту. Виводкові птахи здатні доглядати за собою майже самостійно вже через кілька хвилин після вилуплення. Пташенята багатьох птахів, що гніздяться на землі, таких як фазанові та прибережні птахи, часто здатні бігати практично одразу після вилуплення. На відміну від них, пташенята гніздових птахів безпомічні після вилуплення, сліпі та голі, вони вимагають значних зусиль батьків з догляду за ними. Зокрема, до цієї групи відносяться птахи, що гніздяться в дуплах.
Деякі птахи, такі як голуби, гусаки і японський журавель, створюють пари на все життя і здатні виводити пташенят протягом всього року, без чітко виділеного шлюбного сезону.
Нервова система. Центральна нервова система складається з головного та спинного мозку. Головний мозок досить великих розмірів та є центром вищої нервової діяльності, складається з 5 відділів, характерних для інших хребетних тварин. Півкулі переднього мозку гладенькі, без звивин та порівняно з ссавцями невеликих розмірів. На відміну від останніх складаються переважно не з сірої речовини, а зі смугастих тіл. Півкулі відповідають за управління поведінкою, орієнтацію в просторі, споживання їжі, спаровування та здатність до побудови гнізд. Вища нервова діяльність здійснюється не в неокортексі, як у ссавців, а в гіперстріатумі. Проміжний мозок порівняно невеликий, середній мозок із зоровими буграми. Мозочок розташований у задній частині головного мозку, відповідає за координацію рухів. Довгастий мозок переходить у спинний мозок.
Органи чуття. Основними органами орієнтації птахів є зір та слух. Очі дуже великі за відносними та абсолютними розмірами. Повіки у птахів нерухливі, миготіння здійснюється за допомогою спеціальної перетинки («третя повіка»), яка розміщена в передньому куті ока і рухається горизонтально. Висока гострота зору досягається за рахунок великої кількості фоторецепторів (колбочок та паличок), яких у різних видів на 1 мм3 сітківки припадає від 50 до 300 тис. Іншим важливим пристосуванням є подвійна акомодація (наведення ока на різкість): вона здійснюється шляхом зміни форми кришталика та його одночасним пересуванням м'язами). Поле зору кожного ока становить 150–170°, але поле бінокулярного зору (двома очима) невелике й у більшості птахів становить 20—30°. Лише у сов та деяких соколоподібних очі зсунуті до дзьоба і поле бінокулярного зору зростає. У птахів з добре розвиненим польотом (крячки, ластівки, соколоподібні) на сітківці розташовано 2—3 чутливих плями, які є місцями найгострішого сприйняття. Це обумовлює більшу гостроту зору. Так, сокіл-сапсан може тримати здобич у полі зору на відстані 800–1000 м. Зір у всіх птахів кольоровий.
Орган слуху складається із внутрішнього, середнього та зовнішнього відділів. Внутрішнє та середнє вухо за будовою подібні до вух плазунів, однак канал завитки розвинений більше і відокремлений від мішечка перехватом. У внутрішньому вусі добре розвинений лабіринт, який є органом рівноваги. Слухова кісточка одна — стремінце. Барабанна перетинка занурена нижче за поверхню шкіри та до неї веде канал — зовнішній слуховий прохід, по краю якого у частини видів утворюється складка шкіри — зачаток зовнішнього вуха (добре розвинений у сов).
Нюх у птахів розвинений слабко, смак — досить добре: багато видів розпізнають солодке, солоне, гірке. Смакові бруньки розташовані у слизовій оболонці ротової порожнини, на язиці та при його основі.
Гострий слух та здатність до акустичного аналізу поєднується у птахів із здатністю видавати різноманітні звуки, що несуть біологічно важливу інформацію (звуки спілкування батьків і пташенят, сигнали тривоги, звуки та пози, що регулюють територіальні відносини, взаємовідносини партнерів тощо). Голос і слух забезпечують можливість звукового спілкування не тільки між особинами свого виду, але і між різними видами. Так, наприклад, звук тривоги, виданий сойкою у лісі, сприймається як сигнал небезпеки багатьма птахами. Подібні за зовнішнім виглядом близькі види (наприклад, вівчарики) зазвичай легко розпізнаються за піснею та позивкою. Тому голос відіграє важливу роль ізолюючого механізму, що попереджає міжвидове схрещування.
На відміну від ссавців, у птахів відсутні голосові зв'язки. Основним органом, що видає звуки, є нижня гортань (сиринкс). Звук утворюється внаслідок вібрації тимпанальних мембран (стінок сиринкса) і козелка, що викликаються продуванням повітря через сиринкс. Зміна частоти звуку відбувається завдяки спеціальним м'язам здатним регулювати натяг мембран і діаметр просвіту бронхів.
Вокалізація птахів — набір голосових звуків, які видають птахи. В орнітології поділяється на довгий спів і короткі голосові сигнали, різні за функціями: тоді як спів використовується переважно у статевій поведінці та для захисту території, голосові сигнали призначені переважно для комунікації, наприклад, заклику або повідомлення про небезпеку.
Переважна більшість звуків, що видаються птахами, є мелодійними для людського вуха, що, зокрема, обумовило їх вплив на розвиток музики.
{kind=link}
Самець чорного дрозда під час співу
Визначення
Зазвичай, у вокалізації виділяють спів і голосові сигнали, відмінність між якими засновується на модуляції, довжині та контексті звуків. Спів або пісня зазвичай довший та складніший і асоціюється зі шлюбною та територіальною поведінкою, тоді як голосові сигнали або заклики виконують функції попередження або утримання зграї разом. Інші джерела розрізняють їх засновуючись на функції, у цьому разі короткі звуки, такі як голосові сигнали голубів та невокальні звуки, такі як стук дятлів та змахи крил, вважаються співом. Ще інші автори вимагають від співу складної звукової картини та регулярності у часі, тобто картини, що визначає музику.
Спів найрозвиненіший у птахів ряду горобцеподібних, особливо підгрупи співочих птахів. Здебільшого він видається самцями, а не самками, хоча це не є правилом. Спів найчастіше видається на гнізді, хоча деякі види здатні утворювати його під час польоту. Деякі групи птахів мовчазні, тому що видають лише механічні звуки, наприклад, лелеки лише клацають дзьобами. У деяких манакінів самці розвинули кілька механізмів утворення таких звуків, включаючи стрекотання, характерне для комах. Утворення звуків механічними засобами на відміну від сиринкса, зазвичай, має назву інструментальної музики (за визначенням Чарльза Дарвіна) або механічних звуків та, в роботах сучасніших авторів, сонації. Термін сонація зазвичай визначається як акт утворення невокальних звуків, що утворюються з певним призначенням, та є комунікаційними сигналами, які утворюються неголосовими структурами, такими як дзьоб, крила, хвіст та пір'я.
Анатомія
Голосовим органом птахів є сиринкс. Це кісткова структура в основі трахеї (на відміну від гортані у трахеї ссавців). Сиринкс та інколи повітряні мішки, що оточують його, резонують у відповідь на вібрації, що створюються мембранами, повз які проходить повітря при диханні. Птах контролює частоту звуку зміною напруження мембран. Він контролює як частоту, так і гучність, змінюючи швидкість видоху. Птахи здатні незалежно контролювати обидва боки трахеї, тому деякі види утворюють дві головні частоти одночасно.
Функція
Зазвичай, вважається, що спів птахів розвинувся передусім у результаті статевого відбору як елемент статевої поведінки, зокрема залицяння і привертання самок самцями. Крім того, іншою важливою функцією співу є позначення території. За даними експериментів, якість вокалізації є індикатором пристосованості до умов довкілля. Також, за даними експериментів, на характеристики та частоту співу можуть впливати паразити та хвороби, тож вокалізація є прямим індикатором здоров'я. Репертуар співу також є важливим індикатором пристосованості, здатності самців привертати самок та позначати територію. Часто різні за функцією типи співу продукуються лише протягом певного сезону або у різні частини доби, коли потрібне виконання певної функції, і лише протягом цього часу вони сприймаються іншими птахами. Наприклад, самець західного соловейка продукує спів, призначений для привертання самок, лише уночі (коли співають лише неспаровані самці), а спів, призначений для позначення території, — майже виключно протягом ранкового хору (коли співають всі самці).
Голосові сигнали використовуються передусім для комунікації. Таке спілкування здійснюється як в межах одного виду, так і між видами. Спільні сигнали часто використовуються для привертання індивідуальних птахів до зграї. Ці голосові сигнали характеризуються широким спектром і різким початком і закінченням, а їх повторення, звичайні серед багатьох видів, як вважається, корисні для визначення розташування зграї. Сигнали попередження про небезпеку, на відміну від них, у більшості видів характеризуються високою частотою звука, що утруднює визначення положення птаха, який видає такий сигнал.
Часто птахи можуть дуже добре розрізняти голосові сигнали, що дозволяє їм розпізнавати один одного за голосом. Зокрема, багато птахів, що гніздяться в колоніях, так розпізнають своїх пташенят. Інколи голосові сигнали достатньо відмінні, тому розпізнавати індивідуальних птахів можуть навіть люди, що використовується, наприклад, в екологічних дослідженнях.
Багато птахів здатні подавати сигнали дуетом. Деколи такі дуети настільки синхронізовані, що звучать як один голосовий сигнал. Такі сигнали називають антифонними. Дуетні сигнали відмічені у багатьох родин птахів, включаючи фазанових, саванних сорокопудів, тимелієвих та деяких сов і папуг. Серед сухопутних співочих птахів, птахи найчастіше видають подібні сигнали у разі вторгнення чужаків на їх територію, пропонуючи роль таких сигналів у міжвидовій конкуренції.
Деякі птахи здатні дуже добре імітувати голосові сигнали. У деяких птахів, таких як дронго, імітування сигналів може служити для формування багатовидових зграй.
Деякі печерні види, такі як гуахаро і салангани (роди Collocalia та Aerodramus) використовують звуки слухового діапазону (переважно від 2 до 5 кГц) для ехолокації в темній печері.
Мова та характеристики вокалізації
Мова птахів протягом довгого часу була темою міфів та легенд. Дуже давно було відомо, що голосові сигнали мають певне значення, що відповідним чином інтерпретується слухачами. Свійські кури, наприклад, мають різні сигнали у відповідь на наближення повітряних та наземних хижаків, та відповідним чином відповідають на них. Проте, мова, на додаток до окремих слів, повинна мати і певні граматичні структури та правила. Дослідження подібних структур у птахів є досить важкими через велику кількість можливих інтерпретацій. У одному дослідженні, проте, дослідникам вдалося продемонструвати здатність папуг до утворення граматичних структур, включаючи наявність таких понять як іменник, дієслово та прикметник. Дослідження голосових сигналів шпаків також показали наявність рекурсивних структур.
Зазвичай при описі мови птахів мисливцями та натуралістами виділяють 5 головних типів звуків: заклик, пісня, територіальний сигнал, залицяння і тривога. Перші чотири становлять «базову» поведінку і подаються при відносній безпеці та спокої, тоді як останній означає присутність хижака або іншої загрози. У межах кожної категорії значення звуків залежить від модуляції голосу, руху тіла та контексту.
Слух птахів може виходити за межі слуху людини, опускаючись у деяких видів як нижче 50 Гц, так і вище за 20 кГц, із максимальною чутливістю загалом між 1 і 5 кГц. Діапазон частот голосових сигналів залежить від умов навколишнього середовища, перш за все наявних шумів. Зазвичай вузькі частотні діапазони, низькі частоти, низькочастотна модуляція та довга тривалість звуків та інтервалів між ними характерні для середовищ зі щільною рослинністю (що поглинає та приголомшує звуки), тоді як високі частоти, широкі діапазони, високочастотна модуляція та короткі елементи сигналів характерні для відкритих середовищ. Було також запропоновано теорію, що наявний частотний та часовий діапазон поділяється між різними птахами та їх видами, в результаті при його обмеженні довжина та частотна ширина звукових сигналів зменшуються, цей ефект відомий під назвою «акустичної ніші». Птахи співають голосніше та на більш високих частотах в міських районах, де наявний значний низькочастотний шум.
Діалекти
Вокалізація птахів навіть, одного виду часто досить сильно відрізняється, утворюючи «діалекти». Ці діалекти можуть виникати як через різноманітність довкілля, так і внаслідок генетичного дрейфу, хоча явище мало досліджене, тож вплив окремих факторів залишається невідомим навіть, для добре досліджених видів. Найкраще ці відмінності досліджені для співу під час шлюбного періоду. Проте наслідки цього явища не є однаковими й істотно відрізняються залежно від виду птахів.
Зазвичай самки, що виросли під впливом одного діалекту, не відповідають або відповідають гірше на спів самця того ж виду, що володіє іншим діалектом, що було продемонстровано, наприклад, для білокоронного горобця. З іншого боку, самки, що походять з районів, де поширені кілька діалектів або діалекти різних підвидів, не проявляють такої переваги до одного діалекту.
Подібно до самок була досліджена і відповідь територіальних самців на спів чужаків. Так, зазвичай самці найсильніше відповідають на спів представників власного діалекту, слабкіше на представників свого виду з інших районів, і ще слабкіше на спів споріднених видів, а самці, що поділяють більше пісень з сусідами, краще охороняють свою територію.
У зв'язку з виникненням діалектів часто розглядається питання їх впливу на видоутворення, наприклад, це явище було продемонстроване у роботах з дослідження зябликів Дарвіна. Інші роботи, проте, вказують на суперечливість даних з цього приводу. В результаті загальність результату залишається спірною, а вплив екологічних чинників на цей процес вимагає нових досліджень.
Загальні характеристики
Спів птахів різних видів досить сильно відрізняється один від одного та часто є характерною ознакою виду. Було показано, що саме спів часто є особливістю, що не дає змішуватися представникам споріднених видів, генетично близьких для створення життєздатного потомства. В сучасних дослідженнях спів зазвичай характеризується за допомогою акустичної спектроскопії. Види сильно відрізняються за складністю співу та за числом типів пісень, що може досягати до 3000 у бурого пересмішника; у деяких видів навіть, окремі особини відрізняються за цією характеристикою. У кількох видів, таких як шпаки і пересмішники, спів включає випадкові елементи, запам'ятовані протягом життя птаха у формі мімікрії або «привласнення» (через те, що птах використовує звуки, характерні для інших видів). Ще в 1773 році було встановлено, що в експериментах з вирощування пташенят птахами інших видів, коноплянка була здатна вивчити спів звичайного жайворонка. У багатьох видів здається, що хоча основна пісня і однакова для всіх представників виду, молоді птахи вивчають деякі деталі співу від своїх батьків, при цьому варіації накопичуються, формуючи «діалекти».
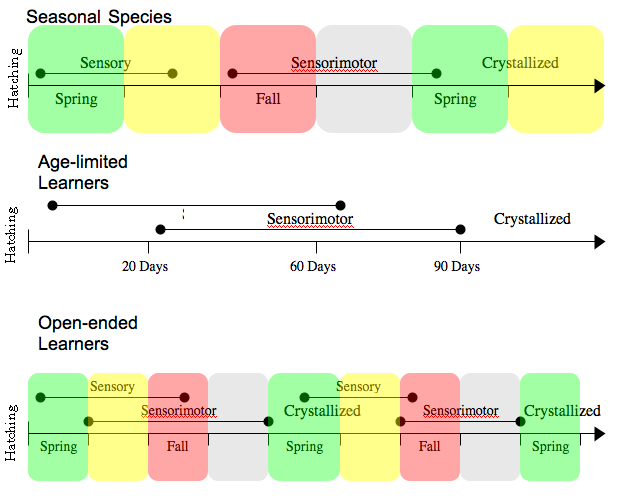{kind=link}
Час навчання співу у різних видів
Зазвичай птахи вивчають пісні протягом початку життя, хоча певні особливості продовжують накопичуватися і пізніше, формуючи спів дорослого птаха. Зяблик-зебра, найпопулярніший модельний організм для дослідження співу птахів, формує спів, що нагадує дорослий, приблизно після 20 діб після вилуплення. До віку біля 35 днів пташеня вже повністю вивчає дорослий спів. Найраніші пісні є досить «пластичними» або підвергнуті змінам, і птаху треба біля 2-3 місяців для доведення співу до остаточної незмінної форми статево зрілих птахів.
Дослідження також вказують на те, що навчання співу є формою моторного навчання, що залучає ділянки базальних гангліїв. Часто моделі навчання птахів використовуються в якості моделей вивчання мови людиною. У деяких видів, таких як зяблик-зебра, навчання обмежене першим роком життя, ці види називають «обмеженими за віком» або «закритими». Інші види, такі як канарки, здатні вивчати нові пісні навіть, у статевозрілому віці, такі види називаються «відкритими» або «необмеженими за віком».
Дослідники висловлювали припущення, що навчання пісень через широке культурне спілкування дозволяє утворенню міжвидових діалектів, що дозволяє птахам адаптуватися до різноманітніших акустичних оточень.
Навчання птахів від своїх батьків було вперше продемонстроване в експериментах Вільяма Торпі 1954 року. Вирощені в ізоляції від самців свого виду птахи здатні співати, а їх спів за загальними рисами зазвичай нагадує спів дорослих птахів, проте не має складних елементів та часто істотно відрізняється. Такий спів часто не здатний привертати самок. Крім співу батьків, пташенятам також важливо чути свій власний спів протягом сенсоромоторного періоду. Птахи, що втратили слух до кристалізації співу, продукують спів, що істотно відрізняється від характерного для даного виду.
Привласнення та імітація
Багато птахів здатні переймати спів не тільки свого виду, але й інших, більш або менш споріднених видів, залежно від птаха. Так, пташенята багатьох видів, вирощені батьками споріднених видів, часто здатні розвивати спів, що нагадує спів прийомних батьків, та в деяких випадках навіть, привертати самок вже цього виду.
Інші птахи здатні переймати спів птахів інших видів, навіть коли вирощуються власними батьками. Кілька сотень видів по всьому світу здатні до такої імітації. Наприклад, назва пересмішник була надана цьому птаху саме за здатність копіювати звуки інших птахів та відтворювати їх у відповідь. Іншим відомим видом, здатним до копіювання, є звичайний шпак, зокрема в Північній Америці, куди цей птах був інтродукований з Європи, він «передражнює» навіть, пересмішника. В Європі та Британії звичайний шпак є найвідомішим імітатором співу інших птахів, що часто відтворює звуки таких птахів як звичайний канюк, звичайна вивільга, великий кульон, сіра сова, качка і гусак. В деяких випадках ці птахи здатні навіть, імітувати голос немовляти або навіть, звуки падаючих бомб під час Другої світової війни. За деякими даними в одному випадку шпак зімітував свисток футбольного судді, чим викликав непорозуміння під час гри.
Лучна очеретянка також до певного ступеня здатна імітувати спів інших птахів, а в її родички чагарникової очеретянки імітація співу інших птахів складає щонайменше 80% всіх продукованих звуків. Деякі дослідники вірять, що ця доля становить 100%, а спірні 20% також були перейняті від недосліджених птахів або навіть, від комах — обидва види очеретянки здійснюють далекі міграції, двічі на рік перетинаючи екватор в Африці, тобто можуть збирати звуки для імітації з величезної території. Новонароджені чагарникові очеретянки починають імітувати навколишні звуки у віці біля 1 місяця після вилуплення, цей процес продовжується протягом міграції та припиняється в січні, коли птаху вже 6-7 місяців, після цього птах втрачає здатність до навчання. За цей період птах встигає запам'ятати спів від 63 до 84 інших видів, з яких в середньому 31 європейський вид і 45 африканських. Деякі з цих африканських видів мають дуже обмежений ареал, через що дослідження співу кожного конкретного птаха дозволяє простежити маршрут його міграції.
Найбільш вражаючим та популярним серед людей прикладом імітації звуків птахами є імітація людської мови. Відомо кілька хвилястих папужок, що виросли у неволі, чий репертуар сягав до 550 слів. Крім того, дуже добрими здатностями для імітації мови людини володіють африканський сірий папуга, австралійські папуги, такі як великий жовточубий какаду і південноамериканські амазони. Олександр фон Гумбольдт під час дослідження Південної Америки описав випадок, коли йому вдалося почути від папуги «мертву мову» знищеного племені. В Європі, де папуги до кінця XV століття були відомі досить мало, здатністю імітувати голос людини були відомі деякі представники родини воронових, такі як галка, сорока і крук.
Проте точні причини такої імітації невідомі. Найімовірнішим є ускладнення власного співу, проте яку користь це може принести птаху, все ще залишається предметом досліджень.
Зрозумілішими є випадки привласнення голосових сигналів, а не співу птахів. Наприклад, товстодзьоба еуфонія часто видає сигнал загрози інших видів під час наближення потенційного хижака до її гнізда, залишаючись сама у безпеці. Подібна поведінка характерна і для звичайної сойки та червоноголової горихвістки. В інших випадках імітація використовується для заманювання жертви у пастку, так панамський чорноспиний лісовий сокіл здатний імітувати заклики про допомогу своїх жертв, а потім ловити птахів, що на них прилетіли.
Нейрофізіологія
В контролі голосових сигналів беруть участь такі відділи мозку:
Шлях формування співу: складається з верхнього вокального центру (high vocal center або hyperstriatum ventralis pars caudalis, HVC), твердого ядра аркопілліума (rubust nucleus of arcopillium, RA) і частини під'язикового ядра, що веде до трахеї та сиринкса (tracheosyringeal nerve, nXIIts);
Передня частина переднього мозку, що відповідає за навчання: складається з латеральної частини магноцелюлярного ядра переднього нового смугастого тіла (lateral part of the magnocellular nucleus of anterior neostriatum, LMAN, гомолога базальних гангліїв ссавців), обасті X (частини базальних гангліїв) і дорсально-латерального відділу середнього таламусу (DLM).
{kind=link}
Шлях навчання співу
Обидва шляхи демонструють статевий диморфізм, зазвичай розвиненіші у самців. Було відмічено, що ін'єкція тестостерону нездатним для співу самкам стимулює ріст HVC і в деяких випадках приводить до виникнення здатності до співу.
Вважається, що утворення співу починається у nucleus uvaeformis таламуса, при цьому сигнали утворюються уздовж шляху, що закінчується на сиринксі. Цей шлях також включає поверхневе ядро nidopallium, HVC, RA, дорсально-латеральний відділ середнього таламусу і нерв трахеї та сиринкса. В літературі також висувалося припущення, що HVC відповідає за утворення складів, тоді як тверде ядро аркопілліума відповідає за чергування складів та формування нот в межах складу.
Під час навчання співу збільшується експресія гену FOXP2, дефекти якого впливають на здатність до розмови та розуміння людини. Також у навчанні грає роль вентральна область покришки (ventral tegmental area, VTA), що посилає сигнал дофаміну до паранюхової долі, області X, LMAN та вентролатераного мозку.
Головним напрямком досліджень нейрофізіологічних аспектів навчання було дослідження методів вивчання співу батьків та зворотний зв'язок з власним співом і його подальша корекція. Експерименти з пошкодження певних відділів мозку виявили такі ефекти:
- Пошкодження шляху формування співу (RA, xXII or HVc) викликає серйозне порушення утворення співу у всіх птахів;
- Пошкодження частин передньої частини передньомозкового шляху, або шляху вокального навчання (DLM і область X) приводить до порушення навчання у всіх птахів;
- Пошкодження LMAN, розташованого у передій частині переднього мозку, у молодих птахів також викликає порушення навчання, але лише у молодих птахів;
- Пошкодження LMAN у дорослих птахів на має ефекту;
- Пошкодження LMAN у дорослих канарок, що характеризуються «відкритим навчанням», викликає порушення співу на всіх етапах життя.
Ці результати показують, що область LMAN є єдиною областю, що характеризується нейропластичністю, а подальші дослідження показали активізацію цієї області у відповідь на власний спів птаха. Ця нейропластичність критична для вивчання нових варіантів співу та необхідна для створення повноти спектру пісень.
Для подальшого дослідження мозкової активності були проведені експерименти зі зміни звуків, які чує птах. За допомогою комп'ютера були відтворені лише певні частини співу або додані окремі звуки, відсутні в співі. В результаті ці дії викликали декристалізацію співу, зокрема втрату як спектральних, так і часових характеристик співу, спів дорослого птаха починав нагадувати спів молодого птаха. Проте при припиненні штучного спотворення співу, птах поступово повертав оригінальні характеристики. Ці результати продемонстрували збереження значної долі пластичності навіть, у птахів із «закритим навчанням», що може бути результатом пластичності області LMAN.
Ідентифікація і систематика
Специфічність вокалізації птахів часто використовується для визначення видів. Зазвичай голосові сигнали описують словами або складами, інколи лінійними діаграмами. Для опису часто використовуються такі слова як крякання, чирикання і щебетання. Ці терміни, проте, сильно залежать від уявлень спостерігача та можуть посилатися на досить різні звуки. Інколи для опису використовуються цілі осмислені фрази, наприклад для опису голосових сигналів смугастої сови англійською мовою часто використовується фраза Who cooks for you? Who cooks for you all? з наголосом на you.
{kind=link}
Використання спектрограм для візуалізації голосових сигналів було впроваджене Вільямом Торпі в 1958 році. Ці візуальні репрезентації зазвичай називаються сонограмами. Багато сучасних визначників птахів використовують сонограми голосових сигналів та співу птахів для ототожнення видів. Сонограма є об’єктивнішим за описові фрази методом документації, проте їх інтерпретація вимагає певного досвіду. Також сонограми можуть бути приблизно перетворені назад на звук.
Визначення видів за співом засновується на спостереженні, що спів у більшості випадків є невід'ємною частиною залицяння і є одним з презиготичних механізмів видоутворення. Багато алопатричних (із різними ареалами) підвидів проявлять відмінності в голосових сигналах та співі. Часто ці відмінності незначні, помітні лише при детальному аналізі сонограм. Проте разом з іншими таксономічними характеристиками подібні відмінності можуть використовуватися для ідентифікації нових видів. Використання вокалізації є причиною розщеплення видових комплексів, наприклад чагарникових жайворонків.
Спів птахів і музика
Серед музикознавців існує думка, що спів птахів значно вплинув на розвиток музики. Силу цього впливу встановити досить важко. Проте відомо, що деякі композитори вставляли спів птахів чи його імітацію у свої твори.
Головними шляхами впливу співу птахів на музику є натхнення під дією співу, свідома імітація співу або пряме вставлення записів співу в музичні твори. Одним з найраніших відомих прикладів імітації співу птахів засобами музичних інструментів є Le Chant Des Oiseaux («Спів птахів») Клемана Жаннекена, написана в XVІ столітті. Іншими прикладами є Sonata Representativa («Показова соната») Генріха Бібера, «Кукушка» Куперена, імітація співу птахів з'являється і в творах великої форми (наприклад у Шостій симфонії Бетховена чи в опері «Зігфрід» Вагнера).
У XX столітті найяскравіше втілив спів птахів у творчості французький композитор Олів'є Мессіан, який цікавився орнітологією. Йому належить 7-частинний «Каталог птахів» для фортепіано, духових і ударних, а також «Пробудження птахів» для оркестру, де імітація пташиного співу лягає в основу музичної тканини творів. Спів птахів міститься і в багатьох інших його творах, наприклад «Квартеті на кінець світу» чи «20 поглядах на немовля Ісуса». Імітація пташиного співу проникла і в джаз, зокрема в твори Flyway («Міграційний маршрут») Пола Вінтера і Grandma Mickey («Бабусін Мікі») Джеффа Сірвербуша.
Ймовірно, першим, хто використав у своєму творі безпосередньо аудіозаписи співу птахів, був італійський композитор Отторіно Респігі у творі Pini di Roma («Римські сосни», 1923–1924). Через кілька років Респігі написав твір Gli Uccelli («Птахи»), заснований на творах стилю Бароко, що містили імітацію співу птахів.
Фінський композитор Ейноюхані Раутаваара в 1972 році написав оркестровий твір Cantus Arcticus, відомий як «Концерт для птахів з оркестром», із значним використанням записів співу птахів Арктики, таких як лебеді, що прилітають туди улітку.
Американський джазовий музикант Ерік Долфі часто слухав птахів перед тим, як практикуватися у грі на флейті. Він стверджував, що в результаті елементи співу птахів входили до складу його імпровізацій.
В психоделічній музиці 1960-х і 1970-х багато рок-гуртів включали спів птахів як звукові ефекти до своїх творів. Англійська група Пінк Флойд включала спів до багатьох пісень своїх альбомів 1969 року Soundtrack from the Film More і Ummagumma. Подібно до них і англійська співачка Кейт Буш включила спів птахів як звуковий ефект до більшості пісень альбому 2005 року Aerial.
Наприкінці 20 століття виник навіть, окремий напрямок музикології — зоомузикологія, присвячений дослідженню музики тварин, засновником якої вважають французького композитора Франсуа-Бернара Маша. Його робота Musique, mythe, nature, ou les Dauphins d'Arion (1983), в якій автор розповідає про музику тварин і зв'язки людини з природою, також включала і розділ з «орнітомузикології».
Політ — головний тип пересування (локомоції) більшості видів птахів, який допомагає їм харчуватися, мігрувати та втікати від хижаків. Вміння літати вимагає від птахів здатності вирішувати чимало завдань, від, власне, польоту до зльоту, посадки та орієнтації; способи вирішення цих завдань сильно залежать від розміру птаха та його екологічної ніші. Хоча певні біологічні адаптації (наприклад, зменшення маси) характерні для всіх птахів, інші (наприклад, форма крил) характерні лише для певних груп та обумовлюють властиві саме їм способи польоту.
{kind=link}
Два звичайних мартини в польоті
Історія дослідження
Початок дослідженням польоту птахів заклав ще Аристотель у роботі «Частини тварин», у четвертій книзі. Він вважав, що швидкість пропорційна силі, яка діє на тіло, тому для його руху постійно необхідний «рушій», що рухає тіло, а сам при цьому залишається нерухомим. Щоб пояснити рух літаючих об'єктів, Аристотель був змушений запровадити поняття передачі функції «рушія» частинам повітря. Поняття інерції, прискорення та аеродинамічного опору тоді ще не були відомі, тому фактично фізика польоту лишилася непоясненою.
Лише через два тисячоліття наступний значний крок уперед зробив Леонардо да Вінчі у своїй праці «Про політ птахів». Його замітки, написані у вигляді інструкцій для птахів, докладно описували, що потрібно робити не тільки при рівномірному польоті, але й для злету й посадки, при пориві вітру та в інших ситуаціях. Його зображення детально показували етапи руху частин тіла птахів. Також він запровадив поняття тиску повітря та його змін навколо крил. Проте роботи Леонардо да Вінчі про політ птахів довго залишалися маловідомими — їх було опубліковано лише в середині ХХ століття.
У праці Джованні Альфонсо Бореллі «Про рух тварин», опублікованій 1680 року, докладно описано анатомію птахів з погляду механіки та збудовано модель, яка пояснювала утворення підйомної сили. Також Бореллі спростував ідею Аристотеля про роль хвоста птахів у регулюванні напрямку польоту.
Наступні етапи розвитку знань про політ птахів пов'язані зі становленням гідродинаміки. Так, Хрістіан Гюйгенс у XVII столітті виміряв залежність аеродинамічного опору від швидкості, а його учень, Готфрід Лейбніц фактично ввів поняття закону збереження енергії. У 1738 році Даніель Бернуллі у роботі «Гідродинаміка» опублікував виведений ним закон, що пов'язував тиск рідини з її швидкістю (зараз відомий як закон Бернуллі), на основі якого Леонард Ейлер вивів набір диференційних рівнянь, що описували рух рідини. Ці рівняння вперше дозволили кількісний опис польоту, хоча і не давали правдоподібних результатів через відсутність у них в'язкості. Лише у 1843 році в роботі Жан-Клода Барре де Сен-Венана, написаній на основі праць Марі-Анрі Нав'є та, незалежно, в роботі 1845 року Габріеля Стокса рівняння Ейлера були доповнені в'язкістю та отримали назву рівнянь Нав'є-Стокса.
{kind=link}
Малюнок польоту птаха, зроблений Леонардо да Вінчі
Перші спроби застосування цих принципів для копіювання польоту птахів і створення літальних апаратів важчих за повітря були здійснені Джорджем Кейлі на початку XIX століття. У своїх працях 1809–1810 років він опублікував перші кількісні розрахунки деталей польоту птахів та вивів форму найменшого опору для заданого об'єму. Він же здійснив перші спроби створення штучних літальних апаратів, які однак, завершилися невдачею. Ці спроби були продовжені Отто Лілієнталем, який також детально досліджував політ птахів і зробив на його основі власний літальний апарат, але його експерименти закінчилися навіть, гірше — сам Лілієнталь загинув від травм, отриманих при аварії. В 1880-их роках Етьєн-Жуль Маре ще далі просунувся у дослідженні польоту птахів, коли зняв перші кінофільми польоту птахів та сконструював дуже складні експериментальні установки для вимірювання сил та тиску повітря в багатьох точках навколо птаха, зокрема він виміряв емпіричну залежність аеродинамічного опору від поверхні.
{kind=link}
Літальний апарат Отто Лілієнталя, 1894 рік
На початку ХХ століття, зі створенням працюючих літаків, основний напрямок гідро- і аеродинаміки змістився з дослідження птахів на дослідження апаратів з нерухомими крилами. Для цих апаратів були створені детальні теорії, і хоча зазвичай вважалося, що їх можна застосовувати і для птахів, експериментальних досліджень практично не проводилося. Великим проривом середини XX століття стало, проте, створення трикутного крила (наприклад, у Конкорда) з метою створення стабільного вихору на передньому краї крила, принцип, що, як було показано пізніше, широко використовується комахами та, ймовірно, птахами.
Лише у 1960-х роках дослідження польоту птахів почалося передусім саме заради вивчення птахів. На той час вже була детально відома функціональна анатомія цих тварин, хоча деякі деталі були відкриті набагато пізніше. Тоді ж стало можливим і використання рентгенівської фотографії для візуалізації кісток в польоті та скорочень окремих м'язів. Також були виміряні витрати енергії на різні цілі при польоті. Дослідження не обмежувалися лабораторними, розвиток радарів дозволив вимірювати швидкість польоту в природних умовах та вивчати стратегію поведінки птахів у різноманітних випадках.
Проте все ще невідомо, наскільки сучасна аеродинамічна теорія може бути застосована до птахів і чи здатна вона описати усі режими їх польоту. Птахи здатні підніматися з місця та приземлятися будь-де, здатні зависати на одному місці та ефективно використовувати вітер. Яким чином вони усе це роблять, досі залишається предметом досліджень.
{kind=link}
Серія фотографій польоту пелікана, зроблена Маре, 1882 рік
Вважається визнаним, що птахи розвилися з невеликих тероподних динозаврів, проте сам механізм виникнення здатності до польоту є одним із найстаріших нерозв'язаних питань палеонтології. Головними є три гіпотези:
«з дерев униз» (англ. arboreal або trees down, Marsh, 1877), за якою предки птахів спочатку навчилися планувати униз із дерев, після чого розвили цю здатність до справжнього польоту за рахунок сили м'язів;
«із землі вгору» (англ. cursorial або ground up, Williston, 1879), за якою предки птахів були невеликими спритними динозаврами, що розвили пір'я для інших потреб, а потім стали використовувати його для підняття у повітря і польоту;
«біг за допомогою крил», варіант «із землі вгору», у якому крила розвилися для утворення направленої униз сили, що уможливлювала кращий контакт з поверхнею, а в результаті — вищу швидкість бігу та здатність бігати по вертикальних поверхнях.
Досі лишається невідомим, чи мав здатність літати перший відомий птах — археоптерикс (Archaeopteryx). З одного боку, археоптерикс мав структури мозку та сенсори внутрішнього вуха, які птахи використовують для контролю свого польоту, а його пір'я було розташоване подібно до пір'я сучасних птахів. З іншого боку, археоптерикс не мав плечового механізму, за допомогою якого сучасні птахи здійснюють швидкі висхідні рухи; це може, зокрема, вказувати на те, що перші птахи не були здатні до махового польоту, але могли ширяти. Наявність більшості викопних решток цього птаха в приморських районах без густої рослинності також призвело до гіпотези, що ці птахи могли використовувати крила для бігу по поверхні води, подібно до ящірок-василісків.
{kind=link}
Археоптерикс, Оксфордський музей орнітології
З дерев униз
Це найперша за часом висловлення гіпотеза, створена за прикладом ширяючих хребетних, таких як вивірки-летяги. За цією гіпотезою, протоптахи, подібні до археоптерикса, використовували кігті для того, щоб підніматися на дерева, з яких вони злітали за допомогою крил.
Однак пізніші дослідження поставили цю гіпотезу під сумнів, наводячи дані, що найперші птахи не вміли лазити по деревах. Сучасні птахи, що мають таку здатність, мають значно скривленіші та міцніші кігті, ніж у тих, що мешкають на землі; кігті ж птахів Мезозойської ери, як і у пов'язаних із ними динозаврів-тероподів, подібні до кігтів сучасних наземних птахів.
Із землі вгору
Пір'я було досить поширеним серед целурозаврів, включаючи раннього тиранозавроїда Dilong, а сучасних птахів палеонтологи найчастіше відносять саме до цієї групи, хоча деякі орнітологи відносять їх до споріднених груп. Функціями цього пір'я могли бути теплоізоляція або ж статева демонстрація. У найпоширенішій версії виникнення польоту «із землі вгору» стверджується, що предки птахів були невеликими наземними хижаками (як сучасний подорожник), що використовували свої передні кінцівки для підтримання рівноваги, а пізніше ці вкриті пір'ям кінцівки розвилися у крила, здатні підтримувати птаха в польоті. Інший варіант гіпотези виводить розвиток польоту зі статевої поведінки та бійок: для привернення уваги протилежної статі розвилося довге пір'я та сильніші кінцівки, що спочатку використовувалися як зброя, а потім опинилися придатними до махового польоту. Також, через те, що багато залишків археоптерикса походять з морських відкладень, було запропоновано, що крила могли допомагати цим птахам рухатися і по поверхні води.
Противники цих гіпотез передусім критикують припущення, що птахи походять саме від целурозаврів. Зокрема, на основі ембріологічного аналізу, що вказує на утворення крил з 2, 3 і 4 пальців (гомологів вказівного, середнього та безіменного пальців людини; перший з цих пальців у птахів формує придаткове крило, для запобігання перекидання в польоті, наприклад при посадці); але передні кінцівки целурозаврів були сформовані пальцями 1, 2 і 3 (великий та наступні два пальці людини). Однак ці ембріологічні дослідження було також піддано сумніву через різні типи розвитку кінцівок у різних груп споріднених тварин, що могли втратити пальці та розвити нові в процесі еволюції.
{kind=link}
Земляна зозуля, що біжить по дорозі, Невада
Біг за допомогою крил
Гіпотеза «бігу за допомогою крил» заснована на спостереженні молодих кекликів та стверджує, що крила отримали свої аеродинамічні функції в результаті потреби швидко бігати по крутих поверхнях, таких як стовбури дерев, або для уникнення хижаків або, навпаки, для несподіваної атаки. Для цього була потрібна сила, що притискає птаха до поверхні. Проте найперші птахи, включаючи археоптерикса, не мали плечового механізму, за допомогою якого сучасні птахи здійснюють швидкі рухи уверх; а через те, що утворення сили униз вимагає цього механізму, ця гіпотеза зазнає значної критики.
Втрата польоту у деяких сучасних птахів
Деякі види птахів, передусім ті, що мешкають на ізольованих островах, де немає наземних хижаків, втратили здатність до польоту. Це доводить, що, незважаючи на великі переваги польоту, він вимагає великих витрат енергії, і тому, коли немає хижаків, може стати непотрібним.
Нелітаючі птахи — птахи, що втратили в процесі еволюції здібність до польоту.
{kind=link}
Пінгвіни (Spheniscus humboldti) під час плавання застосовують крила
Нелітаючі птахи належать до багатьох родин і рядів, тоді як у деяких (пінгвіноподібні, страусоподібні) всі види не літають, в інших (пастушкові) — тільки деякі.
Розміри також коливаються — від 15 см і 35 г у тристанського пастушка до понад 2,5 м і 150 кг у африканського страуса. Деякі вимерлі нелітаючі птахи, такі як епіорніс, були ще більші. Головними фізіологічними особливостями нелітаючих птахів є тонші кістки крил та відсутність або малі розміри кіля — кістки, до якої кріпляться головні м'язи крил (кіль наявний у пінгвінів). Нелітаючі птахи також часто мають більше пір'я.
Багато нелітаючих птахів мешкають в Новій Зеландії — ківі, пастушок такахе, кілька видів пінгвінів, папуга какапо. У багатьох нелітаючих птахів ареал сильно обмежений, як через меншу мобільність, так і через те, що лише на деяких островах, де відсутні наземні хижаки. Проте через обмежений ареал та уразливість до інтродукованих видів, більшість видів нелітаючих птахів знаходиться під загрозою зникнення. Наприклад, так були знищені дронти.
{kind=link}
Какапо, налітаючий папуга
{kind=link}
Пінгвіни, всі види ряду нездатні літати
{kind=link}
Дронт — вимерлий нелітаючий птах
Анатомія структур, пристосованих до польоту
Скелет крила
Передні кінцівки птахів — крила — є головними частинами тіла, пристосованими для польоту. Кожне крило має головну поверхню, якою воно розрізає повітря, що складається з трьох кісток: плечової, ліктьової і променевої. Кисть кінцівки, що еволюційно складалася з п'яти пальців, зредукована до трьох пальців (пальці II, III і IV або I, II, III, залежно від схеми нумерації, а її призначенням є місце кріплення махового пір'я першого порядку, однієї з двох головних груп махового пір'я, що відповідає за форму крила. Другий набір махового пір'я знаходиться позаду кистевого суглобу ліктьової кістки та має назву махового другого порядку. Решта пір'я називається криючим та поділяється на три набори. Інколи крило має рудиментарні кігті, хоча у більшості видів вони втрачаються до часу досягнення птахом статевої зрілості (наприклад, у пташенят гоацинів, що лазять по деревах). Але вони зберігаються у таких птахів як птах-секретар, паламедеї, лапчастоноги, страуси, деякі серпокрильці та у деяких інших видів птахів в якості частої, але не загальної риси. Кігті викопного археоптерикса нагадують за будовою кігті гоацинів.
У гігантських буревісників та альбатросів також існує механізм закріплення суглобів крил в одному положенні для зменшення навантаження на м'язи під час ширяючого польоту.
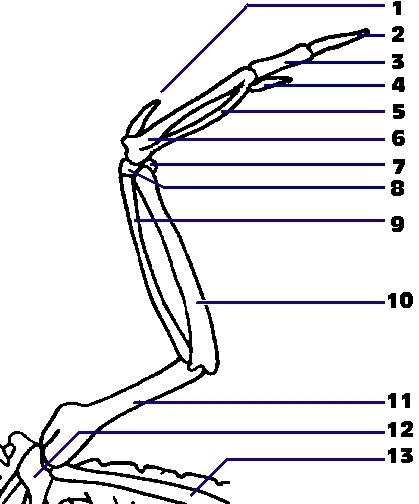{kind=link}
Скелет крила:
1. придаткове крило
2. термінальна фаланга великого пальця
3. базальна фаланга великого пальця
4. малий палець
5-6. карпометакарпус
7. зовнішня ліктьова кістка
8. зовнішня променева кістка
9. променева кістка
10. ліктьова кістка
11. плечова кістка
12. дзьобоподібний відросток
13. лопатка
Пір'я крил
Головним пір'ям, що використовується для польоту та надає крилам та хвосту птахів їх зовнішню форму, є махове пір'я або просто махове. Махове крило зазвичай поділяється на дві або три головні групи: махове першого, другого та інколи третього порядку. Махове першого порядку кріпиться за допомогою сухожилля до кісток кисті, другого порядку — до ліктьової кістки, і третього — до плечової кістки. Здається, для більшості видів птахів махове першого порядку в найбільшій мірі відповідає за здатність літати: навіть повне видалення решти пір'я крил не впливає на дальність та швидкість польоту, але помітне укорочення махових першого порядку, особливо найбільш дистальних, практично позбавляє птахів здатності літати. Велика частина площі махового пір'я вкрита так званим криючим, що захищає його та закриває щілини біля основи.
Махові пера — довге жорстке симетрично розташоване пір’я крил птахів. Головною функцією цих пір'їн є утворення підйомної сили та тяги, надаючи таким чином здатність птахів до польоту. Махові пера деяких птахів також виконують й інші функції, зазвичай пов'язані із статевою поведінкою або живленням. Так, у деяких видів ці пера розвинулися у довге кольорове пір'я, а у інших здатне утворювати звуки в польоті. Маленькі насічки на передньому краю пір'я допомагають совам літати дуже тихо та в результаті успішно полювати. Навіть, нелітаючі птахи мають махові пера, хоча часто у дуже видозміненій формі.
Линяння махових пір'їн часто викликає серйозні проблеми для птахів, зменшуючи їх можливість до польоту. Різні види розвинули різні стратегії запобігання беззахисності протягом цього періоду, від скидання всього пір'я одночасно, зменшуючи період нездатності до польоту, до тривалого періоду линяння, що може займати до кількох років.
{kind=link}
Шуліка рудий (Milvus milvus) в польоті
Махові пера крил
Махові пера крил (лат. remiges) розташовані на задньому (постеріорному) краю крил. Зв'язки прикріпляють їх до кісток, а товстий шар сухожилля забезпечує його жорстке положення. Махові пера птахів симетричні між двома крилами за структурою і формою (окрім випадків мутацій та травм), але не обов'язково за забарвленням. Залежно від розташування, це пір'я поділяється на кілька груп, розташованих уздовж крила.
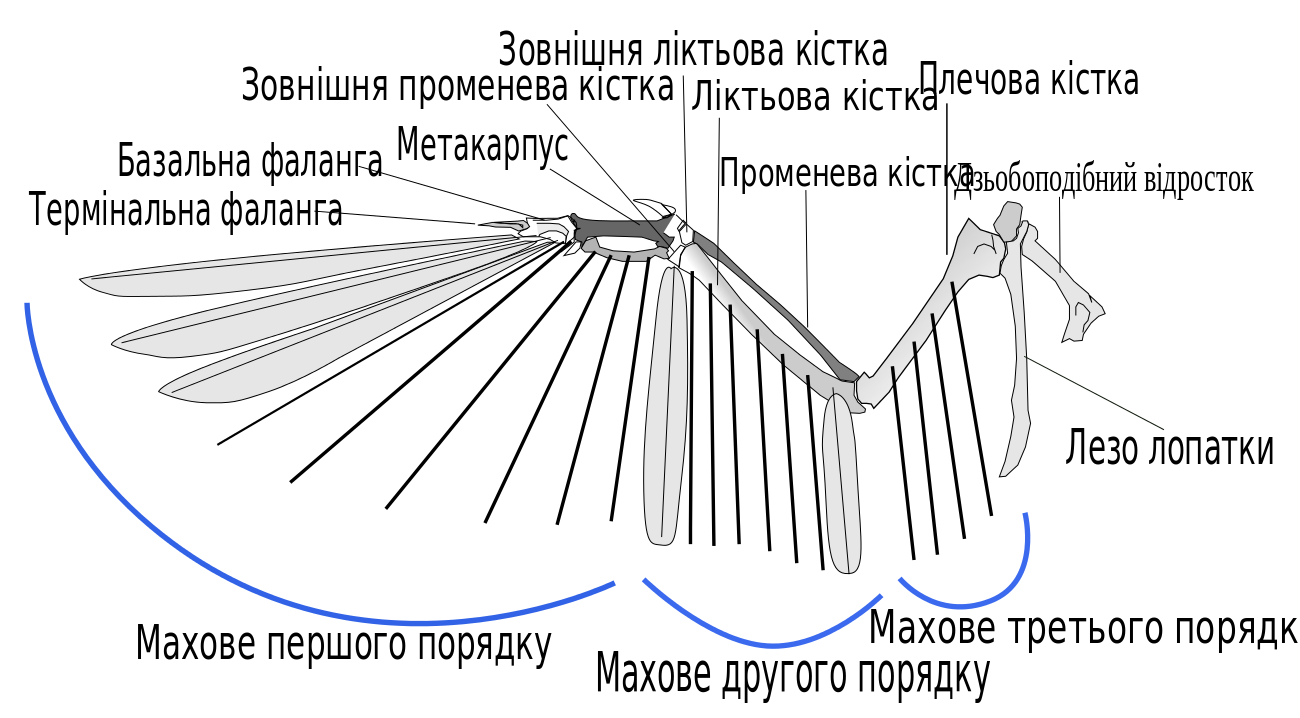{kind=link}
Структура кісток і пір'я крила
Першорядні махові
Першорядні махові пера прикріпляються до кісток мануса («кисті» птахів, що складається з карометакарпуса і фаланги); це найдовше на найбільш вузьке пір'я крил птаха (особливо те, що прикріплене до фаланги), причому кожне перо може незалежно обертатися. Це пір'я особливо важливе для махового польоту та є головним джерелом тяги, просуваючи птаха вперед. Тяга переважно утворюється при русі униз під час махового польоту. При русі уверх (коли крило рухається ближче до тіла птаха) махове першого порядку розходиться та обертається для зменшення аеродинамічного опору, проте надаючи додаткову тягу. Гнучкість пір'я крил також дозволяє великим птахам ширяти при розставленні махового першого порядку для зменшення ефекту утворення вихорів на кінці крила і зменшення аеродинамічного опору.
Різні види птахів істотно розрізняються за числом махових першого порядку. Так, більшість негоробцеподібних мають 9 або 11 на кожному крилі, хоча пірникозові, лелекоподібні і фламінго мають 12, а страуси мають 16. Більшість сучасних горобцеподібних мають 10 махових першого порядку, а деякі 9. Тим, що мають 9, бракує найбільш дистального пера, зазвичай коротшого і часто рудиментарного у горобцеподібних.
Другорядні махові пера
Махові пера другого порядку приєднано до ліктьвої кістки. У деяких видів птахів зв'язки, що сполучають махове крило, приєднані до невеликих проекцій цієї кістки, відомих як пір'єві вузли; у інших видів з’єднання пряме, а цих вузлів не існує. Махові другого порядку залишаються разом протягом польоту, це пір'я нездатне до розведення, як махове першого порядку, та допомагає птаху утворювати підйомну силу через надання аеродинамічної форми крилу. Махові другого порядку зазвичай коротші та ширші за махові першого порядку, із тупішими кінцями. Птахи мають від 6 махових другого порядку у колібрі до 40 у деяких видів альбатросів. Зазвичай більші види з довшими крилами мають більше число цих пір'їн.
Птахи з понад 40 родин негоробцеподібних птахів не мають п'ятого пера на кожному крилі, явище, відоме як діастраксія. До цієї групи відносяться гагари, пірникози, пелікани, яструби, орли, журавлі, баранці, мартини, папуги і сови. У цих птахів п'ятий набір криючого пір'я другого порядку не вкриває махове, ймовірно через поворот пір'я протягом ембріонного розвитку.
{kind=link}
Махові першого (ліворуч) і другого (праворуч) порядку звичайного канюка (Buteo buteo)
Третьорядні махові пера
Махові пера третього порядку прикріплене до плечової кістки та присутні лише у деяких видів. Багато дослідників загалом не використовують термін «махове третього порядку», розглядаючи це пір'я як частину махового другого порядку. Це пір'я діє як захисний покрив для згорнутих махових першого та другого порядку.
Виїмки
Найбільш дистальні махові першого порядку великих птахів, перш за все хижих птахів, часто мають чітко виражені виїмки у різних частинах краю пера. Ці виїмки досить сильно розрізняються за розташуванням та глибиною. Тоді як неглибокі виїмки можуть знаходитися на будь-якій стороні пера, найглибші завжди знаходяться на ширшій задній стороні пера. Ці виїмки можна побачити на зображенні пір’я приблизно посередині обох боків пера, що зображене ліворуч — глибокі на лівій стороні та неглибокі на правій. Функція виїмок все ще залишається спірною. Для її пояснення було запропоновано кілька гіпотез, які, проте, є лише теоретичними, не підкріпленими достатніми експериментальними даними. Зокрема, було запропоновано, що наявність цих виїмок створює щілини на краю крила, що покращує аеродинамічні властивості крила, зокрема збільшуючи підйомну силу, зменшуючи силу аеродинамічного опору та збільшуючи стабільність у польоті.
Придаткове крило
Пір'я придаткового крила зазвичай не вважається маховим — хоча це пір'я асиметричне, воно не має жорсткості, характерної для махового. Проте, пір'я придаткового крила грає важливу роль в польоті, допомагаючи при повільному русі. Це пір'я прикріплене до кісток великого пальця птаха і зазвичай розташоване на передній стороні крила подібно до передкрилків крила літаків, дозволяючи крилу досягати більшого кута атаки та збільшення підйомної сили, запобігаючи втраті швидкості. Рухаючи великий палець та змінюючи щілину між придатковим крилом та рештою крила, птах запобігає втраті швидкості, що необхідно при повільному польоті або приземленні.
{kind=link}
Крижень-самець (Anas platyrhynchos) при посадці, чітко видимі придаткові крила
Затримка розвитку в гоацинів
Розвиток махових пер крил та придаткового крила у гоацинів значно уповільнений в порівнянні з рештою молодих птахів, ймовірно через наявність у молодих гоацинів кігтів на перших двох пальцях. Ці кігті використовуються для лазіння по деревах, а розвиток пір'я заважає цій функціональності. Зазвичай ці птахи відкидають кігті між 70-м і 100-м днями життя, але у деяких вони залишаються і в дорослому стані.
Крильце
Окремою частиною крила є крильце, розташоване на передньому краю крила, біля основи кисті, та здатне розкриватися уперед та вгору від цього краю. За найпоширенішою версією, його функція подібна до функції передкрилків літака, що використовуються для запобігання звалюванню, тобто раптовій втраті швидкості при повільному польоті або гальмуванні при посадці. Інші автори, проте, не погоджуються з цією версією, вказуючи на недостатність даних та пропонуючи гіпотези створення додаткового вихору та стабілізації таким чином початку вихору трикутного крила.
Типові форми крил
Форма крила є важливим фактором, що визначає тип та характеристики польоту, до якого здатний птах. Різні форми крил відповідають різним компромісним рішенням для утворення потрібних характеристик, таких як швидкість польоту, витрати енергії та маневреність. Платформа крила (форма його двовимірної горизонтальної проекції) може бути приблизно описана двома параметрами: відносним подовженням і навантаженням на крило. Відносне подовження крила — це відношення розмаху крил та середньої ширини крила (або квадрат розмаху крил, поділений на площу крил). Навантаження на крило — відношення маси птаха до сумарної площі крил.
Більшість видів птахів можуть бути згруповані за формою крил у кілька загальних типів, хоча цей поділ різниться між різними роботами, а при будь-якому поділі частина птахів займає проміжне положення між ними. Зокрема, звичайним поділом є поділ на еліптичні крила, швидкісні крила, крила з високим відносним подовженням і крила для ширяння, описані нижче.
Еліптичні крила
Еліптичні крила короткі та заокруглені, з невеликим відносним подовженням, що дозволяє птахам маневрувати в обмеженому просторі, наприклад в умовах густої рослинності. Такі крила характерні для лісових хижаків (наприклад, яструба) та багатьох горобцеподібних, особливо неміграторних (міграторні види мають довші крила, потрібні для тривалих перельотів). Також ця форма крил звичайна у видів, що уникають хижаків за рахунок швидкого зльоту, наприклад фазанів і куріпок.
Швидкісні крила
Швидкісні крила короткі та загострені, вони мають високе навантаження на крило та забезпечують високу частоту змахів для надання найбільшої можливої швидкості за рахунок значних витрат енергії. Зазвичай такі крила типові для невеликих птахів та характеризуються дуже великою відносною довжиною кисті крила. Цей тип крил характерний для найшвидших птахів, таких як сапсан, серпокрильці та більшість качок. Подібна форма крил характерна і для алькових, хоч і для іншої мети — альки використовують ці крила для упірнання та «польоту» під водою. Птахи з такими крилами є рекордсменами швидкості — сапсан розвиває швидкість до 140 км/год (за деякими даними навіть, до 300 км/год), а голкохвостий серпокрилець — до 170 км/год.
Особливим випадком крил цього типу є крила колібрі. Їх крила мають найбільшу відому серед птахів довжину кисті, що становить біля 81% довжини всього крила. Ця частина крила у колібрі найбільш рухома, здатна обертатися навколо своєї осі майже на 180°.
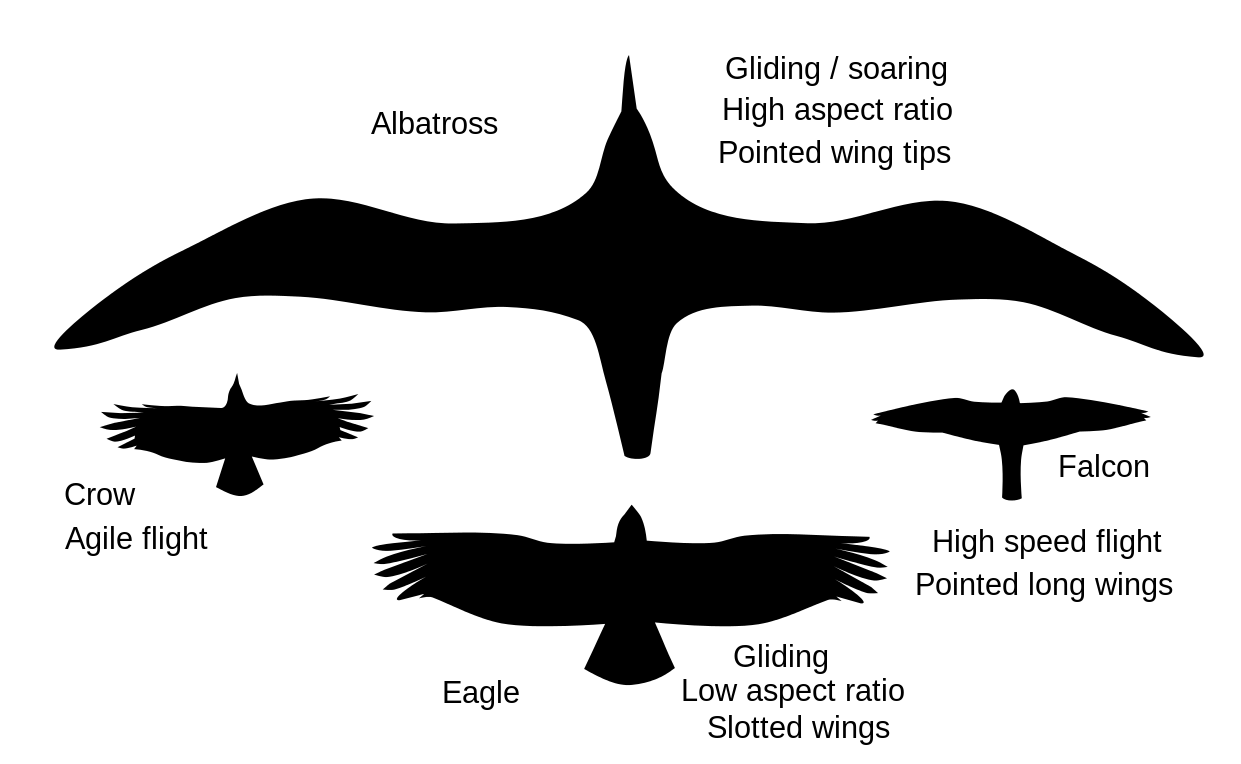{kind=link}
Типові форми крил
Крила з високим відносним подовженням
Крила з високим відносним подовженням є дуже довгими та стрункими, зазвичай вони характеризуються низьким навантаженням на крило та використовуються для повільного польоту, майже ширяння. Такі крила характерні для боривітрів, крячків, дрімлюг та птахів, що здатні переходити на динамічне ширяння над хвилями для збереження енергії, що поширено серед морських птахів.
Особливим випадком цього типу є крила гігантських буревісників та альбатросів, найбільш пристосованих до тривалого ширяння. Ці крила характеризуються найменшою відносною довжиною кисті, біля 25% від довжини крила, та найбільшим числом махових другого порядку — 40. Ці птахи також мають типовий для них механізм закріплення суглобів крил та дещо відмінну будову махового пір'я кисті.
Крила для ширяння
Крила для ширяння досить широкі та характерні для великих сухопутних птахів, таких як орли, грифи, пелікани і лелеки. Глибокі борозенки на кінцях крил між маховим пір'ям зменшують силу опору повітря, а відносно короткий розмір дозволяє підніматися з поверхні землі без потреби у значному розгоні.
Дещо подібні до них крила чапель та ібісів, що зазвичай використовуються для повільного махового польоту.
{kind=link}
Рожевий крячок має крила з низьким навантаженням на крило для повільного польоту, проте здатний пірнати за рибою
Хвіст
В основі хвоста птахів лежать кістки 5-7 хребців та пігостилю, до якого кріпляться махові хвоста. Число махових хвоста різне у різних видів птахів, від нуля у пірникоз до 22-24 у пеліканів, качок та лебедів. Пір'я хвоста зазвичай здатне розкриватися, істотно збільшуючи площу, крім того, хвіст може рухатися в двох напрямках за допомогою шести пар м'язів. Хоча у більшості птахів пір'я хвоста приблизно одної довжини, утворюючи плоский розкритий хвіст, форма хвоста може бути досить різною. Наприклад, у сороки, фазанів, олушів, деяких голубів та зозуль центральне пір'я довше, через що хвіст має округлу форму. У деяких фаетонів, бджолоїдок, поморників, деяких рябків та птахів-секретарів центральне пір'я набагато довше. Навпаки, у сільської ластівки, фрегатів та деяких крячків центральне пір'я набагато коротше, утворюючи вилкоподібний хвіст.
Хвіст бере участь у стабілізації польоту та маневруванні за рахунок як підйомної сили, так і сили тертя. Довгі жорсткі хвости, особливо з довшим центральним пір'ям, найбільш пристосовані для створення тертя, максимально додаючи до стабільності польоту. Навпаки, вилкоподібні хвости утворюють підйомну силу майже без сили тертя, забезпечуючи маневреність при швидкому польоті. Морські птахи часто мають дуже короткі хвости, що нічого не додають до маневреності при повільному польоті, ці птахи можуть дозволити собі такі хвости через відсутність перешкод. Навпаки, у лісових птахів хвости мають надавати їм високу стабільність та стійкість до зіткнень, для чого найзручнішими є довгі відносно прямі хвости.
{kind=link}
Довге центральне пір'я хвоста червонодзьобого фаетона
Інші адаптації до польоту
Хоча утворення крил та пір'я є найпомітнішою адаптацію до польоту, вимоги польоту привели до багатьох інших змін організму птахів.
Скелет птахів досить відмінний від решти сучасних тварин наявністю порожніх кісток, що помітно зменшує його масу. Крім того, у птахів відсутній цілий ряд кісток, що ще існували у археоптерикса, зокрема зник кістковий хвіст, а щелепа з зубами була замінена легким дзьобом. Також в скелеті з'явився кіль, велика кістка, до якої кріпляться махові м'язи крил. Пір'я літаючих птахів також відрізняється від пір'я багатьох нелітаючих наявністю мікроскопічних гачків, що утримують перо разом та надають йому необхідну для польоту міцність.
Важливою адаптацією, призначеною для забезпечення високих витрат енергії та, відповідно, високої швидкості метаболізму, є утворення однонаправленої дихальної системи. Повітря проходить через легені птахів при кожному вдиху та видиху, потрапляючи до повітряних мішків, що дозволяє, на відміну від решти тварин, ефективно використовувати увесь об'єм легенів.
Зліт
Стратегія зльоту може істотно відрізнятися, перш за все залежно від розміру птаха. Маленькі птахи вимагають відносно невеликої або навіть, нульової початкової швидкості, яка генерується за рахунок стрибка. Зокрема така поведінка була продемонстрована на прикладі звичайного шпака та звичайної перепілки, що здатні генерувати 80-90% швидкості польоту за рахунок стрибка, досягаючи прискорення до 48 м/c2. При цьому шпаки часто використовують еластичну енергію гілки, на якій сидять, хоча і не здатні регулювати силу стрибка залежно від її товщини. Інші невеликі птахи, такі як колібрі, чиї ноги занадто тонкі для стрибка, починають махати крилами ще на землі, досягаючи підйомної сили до 1,6 ваги птаха.
Великі птахи зазвичай нездатні злітати з місця і вимагають початкової швидкості для польоту. Найчастіше ця швидкість досягається за рахунок зльоту проти вітру. На додаток часто птахи змушені бігти певну відстань по поверхні землі або води. Деякі великі птахи, такі як орли, використовують скелі, високі гілки дерев або інші високі точки для отримання швидкості за рахунок падіння, морські птахи часто частково здатні досягти подібного ефекту за рахунок зльоту з гребня хвиль.
{kind=link}
Розбіг малого гоголя при зльоті
Посадка
Головним завданням при посадці є зменшення швидкості до безпечного рівня. Основним інструментом для цього у всіх птахів є ноги, на які приземляється птах, що дозволяють як амортизувати удар, так і зберігати вертикальне положення птаха. Проте ефективність амортизації за допомогою ніг сильно варіює між видами. У птахів, що проводять у повітрі більшу частину часу, таких як колібрі, серпокрильці та ластівки, ноги слабкі та не дуже корисні для цієї мети, з іншого боку, у подорожників, тетеруків і куріпок ноги сильні, здатні повністю амортизувати повільний політ цих птахів. Механізм використання ніг дещо варіює від виставлення ніг уперед великими птахами, збільшуючи опір повітря та готуючись до зіткнення з поверхнею, до захоплення гілки, на яку птах збирається приземлитися, невеликими.
На додаток до амортизації ногами, більшість птахів змушені використовувати додаткові механізми. Так, багато птахів, зокрема хижих, завжди приземляються проти вітру. При цьому майже завжди крила розведені убік, а придаткове крило повністю витягнуте. Багато птахів, особливо великих, перед посадкою рухаються нижче за місце посадки (гілки або скелі), а за кілька метрів до цілі піднімаються угору без махання крил, гасячи таким чином швидкість польоту. Цей підхід дозволяє досягати майже нульової швидкості навіть тоді, коли немає вітру.
Гасіння швидкості не так важливе для водоплавних та деяких морських птахів, що, зазвичай, здатні гасити швидкість об воду за допомогою своїх широких ніг. Ноги не обов'язково мають мати перетинки для гальмування — такі птахи, як лелеки, чаплі і журавлі, мають ідеальні для цієї мети ноги. Зазвичай, ці птахи здатні й до посадки на горизонтальну поверхню землі, але часто роблять це незграбно.
{kind=link}
Посадка білоголового орлана
Маховий політ
Не всі птахи і не завжди здатні користуватися різноманітними типами ширяння для отримання енергії, зокрема це майже неможливо для невеликих птахів. Ці птахи змушені отримувати енергію за рахунок змахів крил силою сильних грудних м'язів.
Маховий політ складається з двох окремих типів руху: робочого ходу і зворотного ходу. Під час робочого ходу крило рухається уперед та униз, а зворотний хід повертає крило у початкову позицію. При цьому внутрішня частина крила в першу чергу генерує підйомну силу, тоді як кисть генерує тягу, що штовхає птаха уперед. Під час робочого ходу махове першого порядку зведене разом, формуючи щільну обтічну поверхню. Навпаки, під час зворотного ходу махове першого порядку багатьох, особливо невеликих, птахів обертається навколо своєї осі, дозволяючи протіканню повітря між ним. Більші птахи або довгокрилі маленькі птахи повністю або частково згинають крила, наближаючи їх до тулуба.
{kind=link}
Маховий політ чорного лебедя
Методи збереження енергії
Маховий політ може бути надзвичайно затратним, від 2 до 20 разів перевищуючи основний обмін. Через це птахи були змушені розвити кілька механізмів зменшення витрат при цьому режимі польоту.
Переривистий політ
Одним із засобів збереження енергії під час польоту є переривистий політ, при якому кілька змахів межуються вільним польотом. При цьому існує кілька засобів, якими цей механізм теоретично може зберігати енергію. У великих та середніх птахів під час перерви крила щільно притиснуті до тіла, що дозволяє зменшувати силу тертя. За оцінками, загальна економія енергії при польоті зяблика становить до 35%. Якщо птах тримає крила розкритими під час перерви, політ складається із фази розгону під час махового польоту, а потім планерування під час перерви. Такий політ збільшує підйомну силу та за моделлю дозволяє зберігати до 11% енергії. Здається, птахи можуть користуватися обома стратегіями залежно від швидкості польоту. Крім того, оптимальна частота змахів крил з точки зору використання м'язів може бути більшою за потрібну для польоту, через що переривистий політ надає можливість використовувати м'язи в оптимальнішому режимі.
Переривистий політ украй характерний для деяких груп птахів. Так, він є визначною характеристикою польоту дятлів, що, залежно від виду, махають крилами від 30% до 93% часу польоту, решта приходиться на паузи. Звичайна сорока також проводить до 60% польоту у стані пауз в маханні. Переривистий політ у деяких видів може відбуватися навіть, під час зависання на місці без вітру та з ним, попри це птаху вдається підтримувати своє нерухоме положення відносно землі.
Політ упорядкованою групою
Позаду кінців крил птахів, подібно до крил літаків, утворюються висхідні потоки повітря. Хоча при маховому польоті птахів потоки коливаються, загальна картина така ж сама як й у апаратів з нерухомим крилом. Таким чином, при польоті V-подібною групою кожний птах знаходиться в зоні висхідних потоків попереднього, що дозволяє їм економити енергію. Найбільший ефект досягається для великих птахів, і лише для них переваги перевищують витрати на підтримку точної відстані, через що подібна поведінка найхарактерніша для цієї групи
Зависання
Зависання на місці відносно навколишнього повітря (англ. hovering) є дуже складною задачею, більшість птахів або зовсім нездатні до нього, або здатні це робити протягом дуже незначного проміжку часу. Фактично добре це робити можуть лише колібрі, маса тіла яких варіює між 2 і 8 г. Ненабагато більші нектарниці, що важать від 10 до 20 г, здатні це робити лише якщо немає іншого зручного способу добратися до квітки. При такому зависанні крила колібрі майже невидимі, а крила нектарниць, що рухаються набагато повільніше, видимі як прозорий диск навколо птаха. Під час зависання ці птахи здатні у разі потреби повільно рухатися убік або назад.
При зависанні на місці тіло птаха знаходиться у майже вертикальній позиції, його вісь складає кут 40-50° з горизонтальною площиною. При цьому крила рухаються практично назад-уперед, описуючи вісімку. Як рух уперед так і рух назад здійснюється з деяким пониженням відносно горизонтальної площини, тоді як у крайніх положеннях крила дещо піднімаються угору. Кут руху становить біля 130°, приблизно однаково уперед і назад. Кисть крила при цьому обертається майже на 180° при зміні напрямку руху, постійно утримуючи позитивний кут атаки. Частота руху становить 36-39 Гц у Chlorostilbon lucidus і 27-30 Гц у чорного якобіна (Florisuga fusca), максимальна швидкість — 20 м/с. При цьому колібрі здатні піднімати вантаж вагою 80-200% ваги власного тіла.
{kind=link}
Схема зависання колібрі
Зависання проти вітру
Досить відмінним режимом польоту є зависання відносно поверхні землі за рахунок польоту проти вітру зі швидкістю вітру (англ. windhovering). Така поведінка характерна для багатьох хижих птахів та птахів, що спеціалізуються на полюванні та рибалці: боривітра, скопи, крячків, поморників, деяких рибалочок. Нерухомий стан відносно поверхні допомагає птаху фокусувати очі на поверхні землі та виглядати здобич. Однак цей режим польоту найлегший за оптимальної швидкості вітру, а за іншими умовами вимагає значно більших зусиль.
При зависанні відносно землі голова зазвичай залишається виключно нерухомою. Так, у випадку боривітрів, її коливання складають до 6 мм, причому коливання не корелюють зі змахами крил. При цьому, птаху вдається навіть, достатньо швидко реагувати на зміни швидкості вітру.
Ширяння
Ширянням називається політ без активного використання енергії з боку птаха, що здійснюється або за рахунок втрати швидкості або висоти у спокійному повітрі, або за рахунок використання руху повітря для отримання енергії. Птахи здатні застосовувати кілька стратегій такого використання, описані нижче.
Похиле ширяння
Перешкоди вітру, такі як пагорби, скелі, греблі, дайки, лісосмуги та інші нерівності поверхні змушують повітря у певних місцях навколо них рухатися у вертикальному напрямку. Хоча вимірювання умов повітря над нерівною поверхнею часто є складною задачею, багато видів птахів здатні користуватися такими висхідними потоками, ця стратегія називається похилим ширянням.
Наприклад, при спостереженнях польоту боривітрів над дайкою в Нідерландах було показано, що при вітрі швидкістю 8,7 м/с птахи завжди трималися на висоті 6,5 ± 1,5 м над навітряним боком дайки, підтримуючи кут атаки крила між 6° і 7°.
Динамічне ширяння
Ще одним атмосферним явищем є різниця швидкості вітру на різних відстанях від поверхні, особливо помітна над океаном. При сильному вітрі (7 балів за шкалою Бофорта) його швидкість становить 15 м/с на висоті 10 м, але лише 10 м/с на висоті 1 м. Альбатроси та багато інших морських птахів активно використовують цю різницю, цей процес відомий як динамічне ширяння. Для цього, за поширеною моделлю цього процесу, птах періодично піднімається та опускається, набираючи горизонтальну швидкість у швидших верхніх шарах повітря та вертикальну у нижчих, за рахунок більшої за навколишнє повітря швидкості польоту.
Ширяння над хвилями
Морські птахи також використовують ще два механізми ширяння, пов'язані із наявністю хвиль. Перший тип подібний до описаного вище похилого ширяння та пов'язаний з виникненням висхідних потоків повітря перед хвилею. Так, типова хвиля висотою 1 м і шириною 12 м здатна утворити висхідний потік повітря швидкістю 1,65 м/с. Багато птахів, таких як альбатроси, фульмари, мартини і пелікани, при польоті постійно балансують на навітряній стороні хвилі, користаючись цими потоками. Зазвичай рух відбувається паралельно гребню хвилі. Коли ж хвиля закінчується, птах рухається по інерції, розшукуючи нову хвилю. Рідше, проте, птах може рухатися і разом з хвилею.
Інший засіб отримувати енергію від хвиль полягає у тому, що за хвилею завжди знаходиться ділянка спокійнішого повітря. Птахи, такі як альбатроси і буревісники часто летять там, постійно піднімаючись і опускаючись та фактично користуючись механізмом динамічного ширяння
Термальне ширяння
Ще один тип ширяння пов'язаний з використанням птахами термальних колон — потоків висхідного повітря, що виникають через нагрів повітря від поверхні та нестабільність при підйомі цього повітря угору. Термальні колони найзвичайніші над плоскою поверхнею. Цим методом користуються багато великих птахів, утримуючись в межах колони за рахунок кружляння на місці. Через те, що така поведінка характерна для багатьох хижих птахів, зокрема грифів, шулік, канюків, птахів-секретарів, виникла помилкова думка про очікування птахом здобичі. Цим методом, проте, користуються і лелеки, пелікани та інші птахи, що не є хижими. Птахи, що користуються термальним ширянням, зазвичай мають невелике відносне подовження крила (приблизно 15:1), що робить їх не дуже добрими для швидкісного ширяння, на відміну від, наприклад, альбатросів. Однак таке відносне подовження крила дозволяє кружляти колом меншого радіусу, що допомагає залишатися в межах термальної колони. В деяких випадках термальне ширяння можливе і без кружляння, коли птах летить від одної термальної колони до іншої, набираючи висоту під час прольоту крізь колони.
.jpg){kind=link}
Ширяння чорного шуліки
Маневрування
Маневрування під час польоту досягається за рахунок обертання навколо трьох осей відносно тіла птаха. Обертання навколо осі тіла птаха досягається за рахунок різниці підйомної сили між двома крилами. Птахи з довгими загостреними V-подібними хвостами також допомагають обертанню навколо осі за допомогою хвоста. Обертання навколо горизонтальної осі, перпендикулярної осі птаха, досягається опусканням голови та підняттям хвоста. Третя вісь проходить вертикально, а обертання навколо цієї осі контролюється за допомогою відносної різниці у тязі між двома крилами. Хоча в принципі цей тип обертання може використовуватися для повороту, зазвичай цього не відбувається: замість цього поворот здійснюється за допомогою двох інших типів обертання: птах спочатку обертається навколо осі тіла, а після досягнення певного кута — навколо перпендикулярної осі.
Асиметрична тяга та підйомна сила, необхідні для маневрування, досягаються за рахунок зміни сили тертя і підйомної сили, що здійснюється різним ступенем витягування крил, різним кутом атаки та різною частотою махання крил. Хвіст також допомагає повороту за допомогою нахилу у горизонтальному та вертикальному напрямках. Наприклад, голуби при різкому повороті нахилялися на кут 30°, після чого рухалися по колу в повільному польоті із прискоренням 600 рад/с. Поворот навколо осі птаха в цих експериментах відбувався за рахунок різниці підйомної сили, через те, що махання крилами відбувалося із різницею у частоті в 17%. При цьому зміни кута атаки або площі крил не було зареєстровано.
.
Розмноження
Соціальна організація
Птахи приступають до розмноження у віці від менше 1 року до 12 років. Найбільш раннє статеве дозрівання відзначено у деяких тропічних видів ткачикових (Estrildidae) у 4—5 місяців. Більшість горобцеподібних, багато куроподібних, деякі види качок та голубів починають розмножуватися у віці близько 1 року; багато соколоподібних, сов, серпокрильців, папуг — у віці 2 та 3 років; мартини — від 2 до 7 років; великі трубконоси — до 8—11 років.
Серед птахів 95% видів є моногамними. Вони утворюють пару щонайменше на один гніздовий період, а інколи — на декілька років або до смерті одного з партнерів.
У той же час, серед багатьох видів, які є моногамними, досить звичайною є копуляція з особинами інших пар. Така поведінка є типовою між домінантним самцем та самицею, яка утворила пару з субдомінантним самцем, однак може бути також у результаті примусової копуляції (у качок та інших качкових). Для самиць переваги копуляції з чужим самцем полягають в можливості отримати кращі гени для своїх нащадків та страхування проти можливого безпліддя свого партнера. Самці тих видів, у яких спостерігається копуляція самиць з іншими самцями, уважно охороняють своїх партнерок з метою забезпечення власного батьківства для пташенят, у вирощуванні яких вони беруть участь.
Інші соціальні системи серед птахів включають полігінію, поліандрію, полігамію, полігіноандрію та проміскуїтет. Полігамна система розвивається у тому разі, коли самиця спроможна виростити потомство без допомоги самця. Деякі види можуть застосовувати, залежно від обставин, більше однієї системи.
Гніздування зазвичай включає токову поведінку, яку демонструє, здебільшого, самець. У більшості випадків токова поведінка є досить простою та полягає у різних типах пісень. У той же час деякі види токової поведінки є досить складними. Залежно від виду, вона може включати складні рухи крилами та хвостом, танці, токові польоти або групове токування. У цілому, вибір статевого партнера здійснює самиця, хоча у птахів роду плавунець, які є поліандричними, усе навпаки: простіше забарвлений самець вибирає яскраво забарвлену самицю. Після утворення пари між партнерами може спостерігатися ритуальне годування, «цілування» дзьобами та взаємний догляд за оперенням.
Гомосексуальна поведінка була помічена у самців та самиць багатьох видів птахів, включаючи копуляцію, утворення пари та спільне виховання пташенят.
Територіальна поведінка, гніздування
Практично у всіх птахів у період розмноження добре виражена територіальність — прив'язаність до гніздової території та її захист від вторгнення особин свого, а інколи — й інших видів. Більшість птахів, що гніздиться окремими парами, мають індивідуальну гніздову територію. Такий спосіб дозволяє популяції повніше використовувати придатні для гніздування умови, гнізда таких птахів важче знайти хижакам. У разі колоніального гніздування розмір території, що охороняється, нерідко обмежується найближчим оточенням гнізда. Колоніальність властива птахам, що живляться масовими кормами та збирають їх на деякій відстані від гніздової колонії. Особливо багато колоніальних видів серед морських птахів, що гніздяться на узбережжі (веслоногі, трубконосі, мартинові, алькові). Колоніальність сформувалася під впливом двох факторів: нестачі зручних місць для побудови гнізда та наявність достатньої кількості кормів. Вважають, що перевагами колоніального способу гніздування є успішніший колективний захист від хижаків та ефективне використання трофічних ресурсів.
{kind=link}
Ткачики майстерно роблять гнізда зі стебел рослин
{kind=link}
Глибоке чашоподібне гніздо великої очеретянки
Гніздо. Лише небагато птахів не будують гнізда, відкладаючи яйця безпосередньо на субстрат (дрімлюгові, деякі кулики, кайри). Куроподібні, мартини, сови роблять невелике заглиблення для гнізда у землі з невеликою кількістю вистилки з рослинного матеріалу. Найдосконаліші гнізда роблять дрібні горобцеподібні. Щільне чашоподібне гніздо сплітається з сухої трави та тонких гілочок, а зсередини лоток вистилається м'якими стеблами, мохом, пір'ям та шерстю. У деяких видів гніздо являє собою щільну кулю з товстими стінками та бічним входом (ремез звичайний, синиця довгохвоста). Сільська ластівка робить стінки гнізда з ґрунту, вистилаючи рослинним матеріалом. Дятли довбають дупло у стовбурі дерева, на дно якого без будь-якої вистилки, відкладають яйця.
Гнізда птахи розташовують на землі, на скелястих уступах, розгалуженнях гілок дерев і кущів, заломах очерету. Рибалочки, бджолоїдки, берегові ластівки, топорики риють нори довжиною до 1—2 м. Деякі птахи (лиска, норці, деякі крячки) влаштовують плаваючі гнізда з сухих стебел рослин та водоростей, розташовуючи їх у заростях надводної рослинності. Частина голубів, серпокрильців будують гнізда у порожнинах скель. Багато видів птахів-синантропів влаштовують гнізда у порожнинах різноманітних споруд людини.
Гніздо захищає кладку, насиджуючого птаха та пташенят від ворогів та несприятливих факторів, створює оптимальний температурний режим та вологість.
Пташине гніздо — місце, у якому птах відкладає та висиджує свої, а в деяких випадках чужі яйця і вирощує пташенят. Хоча у популярному розумінні термін зазвичай позначає специфічні, створені самими птахами структури, такі як трав'яні чашоподібні гнізда мандрівного та чорного дроздів, або витончено зіткані підвішені гнізда оропендоли-монтесуми, чорноголового ткачика або червонобривого пардалота, орнітологічне визначення ширше. Для деяких видів гніздом є лише невелике поглиблення в піску, розгалуження двох гілок, нора в землі, дупло у дереві, купа перегниваючого рослинного матеріалу або земляний купол із вхідним тунелем. Деякі види, такі як салангани роду Collocalia, будують гнізда цілком з власної слини, що загусає, утворюючи місце для відкладання яєць на вертикальній стіні печери. Найменші гнізда — гнізда колібрі, що мають вигляд крихітних чашочок діаметром біля 2 см та висотою 2-3 см. З другого боку, гнізда, збудовані джунглевою куркою, досягають понад 11 м в діаметрі та висоти біля 5 м.
Часто сам птах не будує гнізда, захоплюючи чуже гніздо або використовуючи стратегію гніздового паразитизму. В останньому випадку яйця відкладаються у гніздо птаха-хазяїна, того ж або іншого виду, який висиджує яйця та доглядає за пташенятами, позбавляючи птаха-паразита необхідності займатися цим власноруч.
Хоча ступінь участі представників обох статей у будуванні гнізда різна у різних видів птахів, частіше більшу частину роботи виконує самиця. У деяких полігінних видів, проте, більшу частину роботи виконує самець. У деяких видів дорослим птахам у будівництві гнізда допомогають молоді особини від попередніх кладок. Гніздо часто буває місцем різних етапів спаровування, зокрема залицяння і шлюбних ігор, наприклад, такими є гнізда альтаночниць і ткачиків. Здатність будувати та підтримувати добре гніздо ймовірно є результатом еволюційного відбору в цих видів.
Яйце. Птахи розмножуються шляхом відкладання яєць. Пташині яйця мають дуже різні розміри. Із сучасних птахів найбільше яйце в африканського страуса, воно більш ніж в 2000 разів більше за найменше яйце, що належить колібрі. Яйця страуса мають довжину в середньому 180 мм і ширину 140 мм, та важать 1,2 кг. Яйця колібрі 13 мм завдовжки та 8 мм завширшки і важать половину грама. Вимерлий Слоновий птах із Мадагаскару відкладав яйця, що були в 7 разів більші за яйця африканського страуса. У всіх птахів яйця належать до телолицетального типу.
{kind=link}
Порівняльні розміри яєць: яйце африканського страуса праворуч, куряче яйце ліворуч внизу, і перепелині яйця ліворуч вгорі
Зовні яйце вкрите вапняковою шкаралупою, яка на 89—97% складається з СаСО3, 0,5—5,0% — з CaHPO4 та MgHPO4, 2—5% органічної речовини. Тонкий та щільний поверхневий шар шкаралупи називають кутикулою, часто вона блискуча. Кутикула запобігає потраплянню мікроорганізмів усередину яйця через пори в шкаралупі. Шкаралупа забезпечує збереження форми яйця, захист зародку від механічних пошкоджень та висихання, забезпечує газообмін через тонкі звивисті канали у ній, є додатковим джерелом мінеральних солей для формування скелету зародка. Під шкаралупою розташовані дві тонкі підшкаралупові оболонки, які огортають білок. На тупому кінці яйця підшкаралупові оболонки розходяться, утворюючи повітряну камеру. Усередині яйця розташовано білок (білкова оболонка) та яйцеклітина (жовток). Жовток птахів багатий на поживні речовини, які на 85% покривають енергетичні витрати формування зародку. Білок виконує захисну функцію, за рахунок цієї оболонки забезпечується більша частина потреб зародка у воді. Сам жовток являє собою яйцеклітину, на його анімальному полюсі розташовано зародковий диск. Усередині яйця є три зародкові оболонки, що забезпечують життя та розвиток зародка: амніон, алантоїс та хоріон. У більшості видів завдяки відкладанню пігментів, що просочують шкаралупову оболонку, яйця забарвлені. Забарвлення для багатьох видів робить яйця менш помітними.
Усі оболонки яйця формуються під час його проходження яйцепроводом. Повне формування яйця в яйцепроводі триває 5—12 годин, рідко довше. Період часу між відкладанням окремих яєць неоднаковий: у більшості птахів він становить 24 години, у великих птахів — 72 години, а у деяких — до 120 годин (ягнятник).
Розміри яєць у різних видів значно відрізняються не тільки абсолютно, але і відносно величини самого птаха. Як правило, в межах однієї систематичної групи дрібніші птахи відкладають відносно більші яйця. У гніздових птахів яйця дрібніші, ніж у виводкових. У великих кладках яйця відносно менші, ніж у малих. У страусів, великих куроподібних та інших птахів маса яйця становить 1,7—3% маси самиці, у ківі та деяких дрібних куликів маса яйця становить 18—26% маси самиці — це найбільші показники серед птахів.
Кількість яєць, що відкладають птахи, варіює у різних видів. У цілому більший розмір кладки мають виводкові птахи у порівнянні з гніздовими. Одне яйце відкладають трубконосі, великі пінгвіни, більшість чистикових, дрібні колібрі, великі хижаки. Більшість дрібних горобцеподібних частіше мають кладку з 4—6 яєць. У качок частіше 6—10 яєць в кладці. Найбільша кількість яєць — до 22 характерна для куроподібних (сіра куріпка, кеклик).
Стосовно факторів, що визначають величину кладки яєць у птахів висловлено низку гіпотез. Згідно з гіпотезою Д. Лека плодючість виду обмежується здатністю батьків забезпечити їжею потомство, що вони вирощують, і тому величина кладки у гніздових птахів визначається кількістю пташенят, яку вони можуть вигодувати, а у виводкових птахів — кількістю їжі, що вживає самиця перед і під час відкладання яєць.
Усі птахи, за винятком деяких видів, насиджують кладку — обігрівають яйця теплом свого тіла. У насиджуючого птаха на череві і нижній частині грудей випадають пух і частина пер та утворюються оголені ділянки шкіри — насідні плями. Зовсім не насиджують кладку сміттєві кури, до яких належить 11—16 видів. Вони закопують яйця в землю або купу гумусу, де вони розвиваються за рахунок сонячного тепла або тепла, що виникає при гнитті рослин.
Тривалість ембріонального розвитку в межах одного ряду зазвичай менша у дрібніших видів. Найбільша тривалість насиджування — близько 2 місяців — у великих пінгвінів, страусоподібних птахів (страус, казуар, ему, нанду), альбатросів, кондорів та деяких інших. У дрібних горобцеподібних тривалість інкубації — 11—14 діб.
За фізіологічною зрілістю пташенят на час вилуплення птахів поділяють на дві групи: виводкові та гніздові. Пташенята перших вилуплюються добре опушеними, з відкритими очима та слуховими проходами; відразу або невдовзі після вилуплення вони залишають гніздо і можуть самостійно пересуватися й живитися, рухаючись за батьками. Пташенята гніздових птахів вилуплюються голими (веслоногі, папуги, серпокрильці, дятли, горобцеподібні та інші) або слабкоопушеними (частина ракшеподібних та горобцеподібних), з закритими очима та слуховими проходами; у гнізді залишаються, поки не досягнуть майже розміру дорослих і не вкриються пір'ям; потребують регулярного обігріву та вигодовування.
ареалу звичайної зозулі її яйця знаходили в гніздах 300 видів птахів.
Жовток слугує запасом енергетично багатих поживних речовин. Залежно від виду птахів, він складається на 21-36 % з жирів та на 16-22 % з білків (все інше — вода). Жовток підтримується приблизно в центрі яйця двома закрученими жгутами, що називаються халази. Завдяки халазам жовток завжди повернутий таким чином, що зародковий диск знаходиться вгорі, що особливо важливо при насиджуванні яєць птахами. При формуванні жовтка в нього додаються імунні антитіла материнського організму — імуноглобуліни, що представлені переважно імуноглобуліном Y (IgY). Свої власні імуноглобуліни пташеня починає синтезувати через 5-7 днів після виходу з яйця. В ході ембріогенезу зародок поступово обростає та перетравлює жовток, і наприкінці розвитку зародку жовток повністю вміщується в його черевній порожнині. Цим залишковим жовтком, що знаходиться всередині, пташеня продовжує харчуватись ще деякий час після виходу з яйця. Також, в цей час залишковий жовток продовжує постачати імуноглобуліни для імунного захисту організму. У домашньої курки (Gallus domesticus) материнський IgY з залишкового жовтка присутній в організмі курчати протягом перших 14 днів життя, а власний IgY починає синтезуватись починаючи з п'ятого дня. Жовток птахів забарвлений в жовто-оранжевий колір різної інтенсивності. Інтенсивність забарвлення залежить від кількості каротиноїдів, що знаходяться в жовтку і захищають ембріон від хімічного ураження вільними радикалами, які виробляються під час його метаболізму. Таким чином, жовток виконує також і антиоксидазну функцію. Кількість жовтка в яйці може бути істотно різною залежно від виду. Так, новозеландські птахи з роду ківі (Apteryx) не тільки відкладають найбільші серед птахів яйця порівняно із своїм розміром, але й такі, де об'єм жовтка найбільший — понад 50%. У інших видів птахів об'єм жовтка становить від 45-50 % яйця у качок до 15 % у бакланів.
{kind=link}
Структура пташиного яйця
Білок складається з 10 % протеїнів та 90 % води. Він використовується для постачання ембріону вологи, а також виконує роль протиударного захисту та, завдяки великій теплоємності рідини, з якої складається, запобігає раптовим різким перепадам температури навколо ембріону.
Шкаралупа виконує функцію зовнішнього захисту ембріона. Вона вміщує тисячі пор, які забезпечують газообмін. Яйця різних видів птахів можуть дуже розрізнятись за такими ознаками як щільність та товщина шкаралупи. Найтонша шкаралупа у яєць, що відкладають чаплі, в той час як у тропічного птаха Турача (Francolinus francolinus) шкаралупа яєць є найтовщою серед птахів, і становить до 28 % від ваги яйця. Шкаралупа складається з карбонату кальцію СаСО3, який накопичується в організмі самиці птахів протягом всього року, і зберігається, відкладаючись в кістках ніг. У випадку забарвлення шкаралупи вона також виконує функцію камуфляжу.
Шкаралупові оболонки
Із шкаралупою яйця поєднані дві шкаралупові мембрани, внутрішня та зовнішня. Вони захищають яйце від потрапляння всередину бактерій та запобігають надмірному випаровуванню вологи.
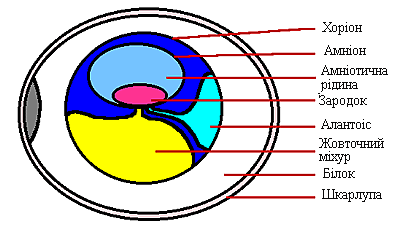{kind=link}
Пташине яйце на початку розвитку зародка
Зародкові оболонки
Всередині яйця знаходяться три зародкові оболонки, що забезпечують життя та розвиток зародка.
Амніон Безпосередньо оточує ембріон, внутрішній шар клітин амніону продукує амніотичну рідину, в якій плаває зародок. Ця рідина підтримує оптимальний для покровів ембріона склад іонів та захищає його.
Хоріон Оточує всі ембріональні структури і слугує захисною селективною мембраною, що виконує функції газообміну.
Алантоїс (або алантоісний міхур) росте разом з ембріоном, зростається з хоріоном, утворюючи хоріо-алантоісну мембрану. Разом з хоріоном забезпечує дихання (обмін кисню та вуглекислоти) та виділення. Алантоіс є вмістом азотистих відходів, що продукують нирки ембріону під час його розвитку (сечова кислота).
Всі перераховані оболонки разом з зародком спочатку займають дуже невеликий об'єм яйця, формуючи зародкову бляшку на поверхні жовтка. По мірі розвитку зародок та його зовнішні оболонки ростуть, поступово утилізуючи весь вміст яйця. При виході пташеняти з яйця зародкові оболонки відриваються і залишаються в пустій шкаралупі.
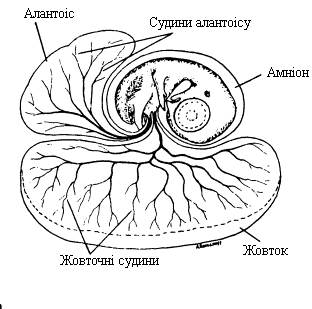{kind=link}
Семиденний курячий ембріон
Забарвлення
Звичайно однотонно-білі яйця несуть птахи, які гніздяться у норах, печерах або дуплах — ластівки берегові, рибалочка, дятли, крутиголовки та ін. Яскраво-білий колір яєць при цьому допомагає птахам легше знаходити їх в затемненому середовищі. Птахи, які гніздяться на землі або у відкритих гніздах на деревах, частіше за все мають забарвлені яйця (однотонні або плямисті) — такий камуфляж допомагає уникати хижаків. Втім, деякі види, що гніздяться на землі, відкладають білі яйця — але в цьому випадку яйця швидко набувають кольорів навколишнього середовища, вкриваючись плямами від ґрунту та гниючих рослинних рештків у гнізді. У деяких випадках, як, наприклад, у тонкодзьобої кайри (Uria aalge), кожна самиця відкладає яйця іншого забарвлення, що допомагає їм знаходити свою кладку серед тисяч таких самих всередині пташиного базару, на скельних полицях. У іншого виду, Індійської жовтосережкової чайки (Vanellus malabaricus) яйця мають різне забарвлення залежно від субстрату, на якому вона гніздиться.
Форма
Основні форми яєць можуть бути такими: кулеподібні, як у дятлів та сов; звужені з одного кінця — у деяких птахів це є способом компактного розміщення яєць в гнізді вузькими кінцями всередину, що дає можливість невеликому птаху одночасно і рівномірно обігрівати всі яйця (наприклад, сивки), а у інших птахів така форма яєць є захистом від скочування з скельних полиць (кайри); видовжені вузькі яйця характерні для птахів з вузьким тазом, що відкладають яйця відносно великого об'єму — це є анатомічним пристосуванням; і, нарешті, звичайні яйця овальної форми, наприклад курячі.
Наука, що вивчає пташині яйця, називається оологія.
Гніздовий паразитизм
Це явище характерне для деяких видів птахів, які не роблять власних гнізд, не насиджують кладку та не виводять пташенят, а підкладають яйця в гнізда інших птахів, які насиджують ці яйця та вигодовують чужих пташенят. Гніздовий паразитизм характерний для близько 80 видів птахів, що належать до 4 рядів: 40 видів зозуль, 6 видів трупіалів (Icteridae), 6 видів медоуказчиків (Indicatoridae), 3 видів африканських ткачиків та чорноголової деревної качки (Heteronetta atricapilla). У звичайної зозулі період яйцекладки триває 35—45 днів, за цей час самиця з інтервалом 1—3 доби відкладає 12—20 яєць. Кожна самиця відкладає яйця певного забарвлення, але забарвлення яєць різних самиць варіює в широких межах. У межах ареалу звичайної зозулі її яйця знаходили в гніздах 300 видів птахів.
Міграція, або переліт птахів — щорічне переміщення або переселення птахів на відносно великі відстані, пов'язане із сезонною зміною екологічних або кормових умов або особливостями розмноження з території гніздування до території зимівлі і назад, одна з форм міграції тварин. Часто у визначенні також ставиться вимога здатності до такого переміщення у відповідь на довжину світлового дня або час року, незалежно від погодних умов, бо тільки ці фактори здатні забезпечити точну періодичність. Міграція — пристосування до сезонних змін клімату та залежних від них факторів (наявність корму, відкритої води тощо). Здібності птахів до міграції сприяє їхня висока мобільність завдяки здатності до польоту, недоступна більшості інших видів наземних тварин.
Типи переміщень птахів
За характером сезонних переміщень птахів поділяють на три основні категорії: осілих (що постійно мешкають на відносно невеликій території), кочові (що здійснюють переміщення на відносно значні відстані не регулярно, лише у пошуках їжі або у випадку поганої погоди) і мігруючих або перелітних (що здійснюють більш-менш далекі сезонні переміщення). Проте цей поділ вельми умовний, як через існування безперервного спектру моделей поведінки між цими категоріями, так і через те, що в межах однієї популяції птахи можуть демонструвати різну поведінку, а один конкретний птах — змінювати її в окремих випадках протягом життя.
Крім того, за певних умов, птахи, як і інші тварини, можуть виселятися з певної території без повернення назад або інвазуватися (впроваджуватися) в регіони за межами району їхнього постійного проживання; такі переселення безпосередньо до міграції не відносяться. Виселення або впровадження може бути пов'язане з природною зміною ландшафту — лісовими пожежами, вирубкою лісів, осушенням боліт і т. д., або з перенаселенням конкретного виду на обмеженій території. У таких умовах птахи вимушені шукати собі нове місце, і таке переміщення ніяк не пов'язане з їхнім способом життя або часом року. До впроваджень також часто відносять інтродукцію — навмисне переселення видів у регіони, де вони ніколи раніше не мешкали.
Як вже зазначалося, дуже часто не можна однозначно сказати, що даний вид птахів є строго осілим, кочівним або перелітним: різні популяції одного і того ж виду, і навіть, птахи однієї популяції, можуть поводитися по-різному. Наприклад, кропив'янка (Sylvia) на більшій частині ареалу, включаючи майже всю Європу і приполярні Командорські і Алеутські острови, живе осіло, у Канаді і на півночі США кочує на незначні відстані, а на північному заході Росії, в Скандинавії і на Далекому Сході є перелітним птахом. У звичайного шпака (Sturnus vulgaris) або блакитної сойки (Cyanocitta cristata) можлива ситуація, коли на одній і тій же території частина птахів у зимовий час переміщується на південь, частина прибуває з півночі, а частина живе осіло.
Більшість міграцій відбуваються широким фронтом, проте в деяких випадках вони відбуваються вузькими смугами, міграційними маршрутами. Зазвичай подібні маршрути йдуть уздовж гірських хребтів або прибережної смуги, що дозволяє птахам користатися висхідними потоками повітря аби запобігти подолання географічних бар'єрів, таких як довгі простори відкритого моря. Також, маршрути не обов'язково збігаються в обох напрямках перельоту.
Більшість великих птахів перелітають зграями, що часто формують регулярні розташування птахів, такі як V-подібний «клин» з 12—20 птахів. Таке розташування допомагає птахам зменшити енергетичні витрати на переліт. Наприклад, ісландський (Calidris canutus) і чорногрудий побережники (Calidris alpina), як було визначено за допомогою радарів, летять у зграях на 5 км/год швидше.
Висота польоту також різна у різних видів птахів. Так, залишки шилохвостів (Anas acuta) і великого грицика (Limosa limosa) були знайдені під час експедиції на Еверест на висоті до 5 тис. м у льодовику Хумбу. Гірські гуски (Anser indicus) спостерігалися під час перельоту над вершинами Гімалаїв на висоті біля 8 тис. м, навіть коли неподалік були низькі перевали висотою 3 тис. м. Морські птахи зазвичай летять дуже низько над морем, проте піднімаються при прольоті над землею. У сухопутних птахів спостерігається зворотна картина. Проте, більшість міграційних птахів летять на висоті від 150 до 600 м. Зіштовхнення птахів з літаками зазвичай відбуваються на висоті до 600 м і майже ніколи вище 1800 м.
Не всі птахи здійснюють міграції за допомогою перельоту. Більшість видів пінгвінів (Spheniscidae) здійснюють регулярні міграції вплав, шляхи цих міграцій можуть досягати 1000 км довжиною. Блакитний тетерук (Dendragapus obscurus) здійснює регулярні міграції на різні висоти переважно пішки. Протягом засухи довгі міграції пішки здійснюють і австралійські ему (Dromaius).
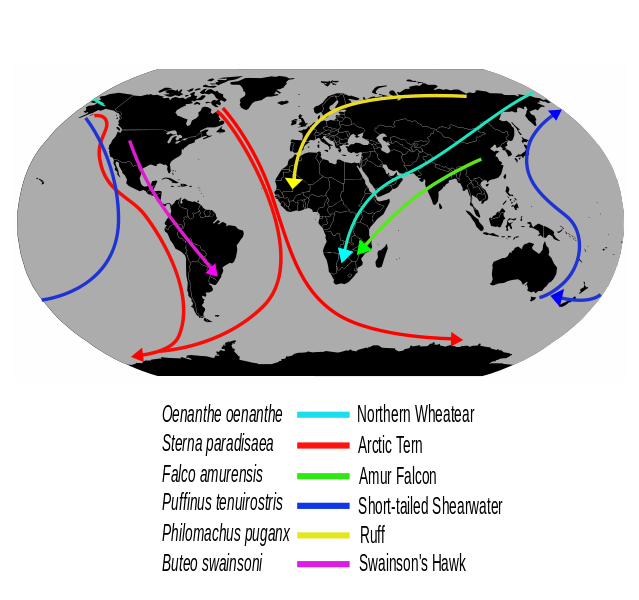{kind=link}
Приклади маршрутів міграції
Осілі птахи — птахи, що не здійснюють регулярних міграцій, у протилежність до міграційних птахів.
Багато популяцій птахів залишаються на одному місці за сприятливих природних умов, проте за нетипових природних умов (наприклад, виключно холодної зими або посухи) або зміни чисельності популяції ці птахи можуть здійснити міграцію до іншого району.
Переважна більшість видів осілих птахів мешкають в таких умовах, де сезонні зміни не впливають на доступність корму — тропічному і субтропічному кліматі. У помірному і північному поясі таких птахів небагато; до них зокрема відносяться синантропи — птахи, що мешкають поблизу людини і залежні від неї: сизий голуб (Columba livia), хатній горобець (Passer domesticus), сіра ворона (Corvus cornix), галка (Corvus monedula) і деякі інші. Частина осілих птахів, яких також називають напівосілими, поза сезоном розмноження переміщається на незначні відстані від своїх місць шніздувань — на території України до таких птахів можна віднести, зокрема, глушців (Tetrao urogallus), рябчиків (Bonasa bonasia), тетеруків (Tetrao tetrix), частково сорок (Pica pica) і звичайну вівсянку (Emberiza citrinella).
Міграція на великі відстані
Типова картина міграції північних сухопутних птахів, таких як ластівки (Hirundo) і хижі птахи, являє собою міграцію до тропічних регіонів. Багато качок, гусок (Anser) і лебедів (Cygnus) північної півкулі є перелітними птахами, проте вони мігрують лише настільки, наскільки необхідно уникнути замерзлих водойм своїх північних районів гніздування. Більшість видів мисливських птахів залишаються в північній півкулі, але в районах із м'якішим кліматом. Наприклад, короткодзьобий гуменник (Anser brachyrhynchus) мігрує з Ісландії до Британії та прилеглих районів. Шляхи міграції та райони зимування зазвичай вивчаються молодими птахами під час першої міграції разом із своїми батьками. Деякі інші качки, проте, такі як велика чирянка (Anas querquedula), повністю або частково переміщаються до тропіків.
Природні бар'єри мають подібну роль і для морських птахів, але зворотну в порівнянні з сухопутними: значні безводні райони, де неможливо прохарчуватися, є для них непереборними бар'єрами. Відкрите море також може бути бар'єром для птаха, що звик харчуватися у прибережних водах. Для запобігання перешкод птахи часто змушені робити перельоти обхідними шляхами: наприклад, чорна казарка (Branta bernicla) мігрує від півострова Таймир до Ваденського моря через узбережжя Білого і Балтійського морів, замість прямого перельоту через Північний Льодовитий океан та північну Скандинавію.
Подібна ситуація спостерігається і в прибережних птахів. Багато видів, такі як чорногрудий (Calidris alpina) і американський побережники (Calidris mauri), здійснюють довгі міграції зі своїх арктичних районів гніздування до тепліших районів тієї ж півкулі, інші, такі як довгопалий побережник (Calidris pusilla), подорожують до тропіків. Прибережні птахи, як і великі водоплавні птахи, характеризуються значною витривалістю в польоті. Це дозволяє їм у випадку зимування в помірних районах здійснювати подальші короткі перельоти у випадку несприятливої погоди.
Для деяких з прибережних птахів можливості перельоту залежать від наявності певних видів харчових ресурсів у ключових місцях зупинки уздовж шляху міграції. Це дозволяє цим птахам отримати достатньо їжі для наступної ділянки подорожі. Наприклад, важливими місцями зупинки багатьох видів птахів є затока Фанді і Делаверська затока.
Найбільшу відстань без зупинки серед всіх перелітних птахів здатні пролітати деякі популяції малого грицика (Limosa lapponica), що перелітає більше 11 тис. км з арктичної тундри Алеутських островів до новозеландських районів зимування без зупинок. Перед початком перельоту жир становить 55% маси тіла, що необхідно для забезпечення енергією такої довгої подорожі.
Картини міграції морських птахів подібні картинам водоплавних і прибережних птахів. Деякі птахи, такі як чорний чистик (Cepphus grylle) і деякі мартини (Larinae), досить осілі, інші, такі як більшість крячків (Sterna) і гагарок (Alcidae), гніздяться в помірних районах північної півкулі та перелітають різні відстані для зимування. Полярний крячок (Sterna paradisaea) здійснює найдовші міграції з усіх птахів, що дозволяє йому отримувати більше сонячного світла, ніж будь-якому іншому птаху, через те, що він мігрує від арктичних районів гніздування до антарктичних районів зимування. Один з полярних крачків, окільцьований ще пташеням на островах Фарне біля східного узбережжя Великої Британії, досяг Мельбурна (Австралія) вже через три місяця після вилуплення, здійснивши мандрівку довжиною більш ніж у 22 тис. км. Кілька видів морських птахів, зокрема океанник Вільсона (Oceanites oceanicus) і великий буревісник (Puffinus gravis), гніздяться в південній півкулі та мігрують на північ протягом південної зими. Ці морські птахи мають перевагу над більшістю перелітних птахів оскільки здатні знаходити їжу протягом польоту над відкритим океаном.
Більшість морських птахів, особливо представників ряду буревісникоподібних (Procellariiformes), перелітають на значні відстані, зокрема альбатроси (Diomedeidae) південних океанів можуть перелітати всю планету поза сезоном гніздування. Ці птахи широко поширені над всім океаном, хоча концентруються в районах, де знаходять найбільшу кількість їжі. Багато з них наближаються до рекордів довжини перельоту, так сірий буревісник (Puffinus griseus), що гніздиться на Фолклендських островах, мігрує на відстань близько 14 тис. км від районів гніздування до районів Північного Льодовитого океану біля Норвегії. Деякі менські буревісники (Puffinus puffinus) здійснюють таку ж подорож у зворотному напрямку. Оскільки ці птахи відносно довгоживучі, за життя вони долають величезні відстані, за підрахунками один рекордний менський буревісник пролетів 8 млн. км за 50 років життя.
Деякі птахи з великими крилами залежать від термальних колон висхідного теплого повітря, що дозволяють їм ширяти. Ці птахи включають багатьох хижих птахів, таких як грифи, орли і канюки, та деяких інших, наприклад, лелеки (Ciconia). Ці птахи мігрують протягом світлої частини доби. Перелітні представники цієї групи зазвичай не здатні долати великі водойми через відсутність над водою термальних колон та нездатність летіти безперервно протягом тривалого часу. Середземне море, як й інші моря, є для них майже непереборним бар'єром, що змушує птахів знаходити вузькі місця або обхідні маршрути. Велика кількість хижих птахів і лелек перетинає моря в районі Гібралтарської протоки, протоки Ересунн та протоки Босфор під час перельоту. Деякі численні види, такі як звичайний осоїд (Pernis apivorus), перелітають через ці протоки в кількості сотень тисяч за один сезон. Інші бар'єри, такі як гірські хребти, також викликають концентрацію птахів у районі вузьких проходів, особливо великих денних птахів. Це дуже помітно при перетинанні птахами Центральної Америки.
Багато малих комахоїдних птахів, зокрема горобцеподібні (Passeriformes), колібрі (Trochilidae) і мухоловки (Muscicapidae), також перелітають значні відстані, переважно вночі. Вони приземляються вранці і часто роблять зупинку на кілька днів перед продовженням перельоту. Ці птахи часто називаються транзитними мешканцями в районах, де вони мешкають протягом короткого часу між початком та закінченням перельоту.
За допомогою перельоту вночі, нічні перелітні птахи зменшують загрозу від хижаків та запобігають перегріву від збільшених витрат енергії для перельоту. Також це надає їм можливість харчуватися протягом дня щоб забезпечити себе енергією для польоту. Недоліком такої поведінки є неможливість достатньо висипатися. Перелітні птахи, здається, здатні змінювати свою потребу в сні для компенсації втрат.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Кочівля та міграція на короткі відстані
Кочівними називають птахів, які поза сезоном розмноження пересуваються з місця на місце у пошуках їжі. Такі пересування зазвичай не пов'язані з циклічністю і повністю залежать від доступності їжі та погодних умов, у такому випадку вони не вважаються міграцією. Проте, існує цілий спектр проміжних між кочівлею і довгою міграцією моделей поведінки птахів, зокрема коротка міграція, що викликається безпосередньо погодними та харчовими умовами і має відносно регулярний характер. Проте, на відміну від довгої міграції, птахи значно змінюють час відправлення до подорожі залежно від погодних умов та можуть пропускати міграцію в теплі або іншим чином сприятливі роки.
Наприклад, мешканці гір та боліт, такі як червонокрилий стінолаз (Tichodroma muraria) і пронурок (Cinclus cinclus) можуть пересуватися лише на різні висоти, уникаючи холодної гірської зими. Інші види, такі як кречет (Falco rusticolus) і жайворонок (Alauda), переміщаються до узбережжя або у південніші райони. Інші, такі як зяблик (Fringilla coelebs), не міграційні в Британії, але перелітають на південь з Ірландії в дуже холодну погоду.
Кочівні птахи ряду горобцеподібних мають два типи еволюційного походження цієї поведінки. Ті види, що мають родичів, перелітних на великі відстані, в тій же родині, такі як вівчарик-ковалик (Phylloscopus collybita) є видами, що походять з південної півкулі, які поступово скоротили довжину зворотного перельоту таким чином, що назавжди залишилися в північній півкулі. На відміну від них, види, які не мають ніяких міграційних родичів, наприклад омелюх (Bombycilla), фактично перелітають у відповідь на зимову погоду, а не заради збільшення можливостей відтворення.
У тропіках існує незначна варіація в довжині дня протягом всього року і завжди достатньо тепло для адекватного харчування. На відміну від сезонних рухів для зимування птахів помірних широт, більшість видів є в широкому сенсі осілими. Проте, багато видів перелітають різні відстані залежно від кількості опадів. Так, багато тропічних районів мають вологі та сухі сезони, найкращим прикладом яких є мусони Південної Азії. Прикладом птаха, який перелітає залежно від кількості опадів, є лісовий зимородок (Halcyon senegalensis), мешканець Західної Африки. Існує кілька видів, виключно зозуль, які є дійсними перелітними птахами в межах тропіків. Прикладом є мала зозуля (Cuculus poliocephalus), яка гніздиться в Індії та проводить решту часу в Африці.
У високих горах, таких як Гімалаї і Анди, багато видів здійснюють також сезонні висотні переміщення, а інші можуть здійснювати довгі міграції. Так, гімалайські кашмірська мухоловка (Ficedula subrubra) і строкатий дрізд (Zoothera wardii) можуть переміщатися на південь до плоскогір'їв Шрі-Ланки.
Довга міграція птахів є переважно, хоча і не виключно, явищем північної півкулі. У південній півкулі сезонні міграції менш помітні. На те є декілька причин. По-перше, значні безперервні простори суші або океану не викликають звуження міграційних маршрутів, що робить міграцію менш помітною для людини-спостерігача. По-друге, щонайменше на суші, кліматичні зони зазвичай поступово переходять одна в одну, не створюючи різких стрибків: це означає, що замість довгих перельотів над несприятливими районами для досягнення певного місця, міграційні птахи можуть мігрувати повільно, харчуючись протягом подорожі. Часто без спеціальних досліджень непомітно, що птахи в певному районі мігрують, бо різні члени того ж самого виду прибувають протягом різних сезонів, поступово рухаючись у певному напрямку.
Проте, багато видів дійсно гніздяться в помірних районах південної півкулі та зимують у північніших тропічних районах. Наприклад, такі міграції здійснюють південноафриканська велика смугаста ластівка (Hirundo cucullata) і австралійські шовкова міагра (Myiagra cyanoleuca), австралійський широкорот (Eurystomus orientalis) і райдужна бджолоїдка (Merops ornatus).
Вторгнення і розосередження
У деяких випадках умови навколишнього середовища, наприклад, різке зниження кількості їжі після багатого на їжу періоду, приводить до вторгнення птахів в інші райони, коли велика кількість птахів разом залишають звичайні райони мешкання. Звичайний омелюх (Bombycilla garrulus) та ялиновий шишкар (Loxia curvirostra) є прикладами видів, що проявляють значні варіації чисельності птахів щороку, і таким чином сприятливі до впроваджень.
Помірні зони південних континентів мають значні посушливі райони, особливо в Австралії та південно-західній Африці, де міграції, викликані погодними умовами, звичайні, але не завжди передбачувані. Кілька тижнів дощів в одній або іншій частині зазвичай сухої центральної Австралії, наприклад, викликає швидкий ріст рослин та безхребетних, що харчуються ними, привертаючи птахів з суміжних районів. Це може трапитися протягом будь-якої частини року, та, в кожному конкретному районі, повторюється не частіше ніж раз у десятиліття, залежно від частоти періодів Ель-Ніньйо і Ла-Нінья.
Фізіологія і контроль
Контроль часу міграції та факторів навколишнього середовища, у відповідь на які відбувається міграція, закодовані на генетичному рівні та до деякого ступеня проявляються навіть, у багатьох осілих видів птахів. Здатність до навігації і орієнтації протягом міграції є набагато складнішим явищем, яке може включати як генетичну інформацію, так і навчання.
Час міграції
Головним фізіологічним фактором, що впливає на вибір часу міграції, є зміни довжини світлового дня. Ці зміни також пов'язані з гормональними змінами організму птахів.
Безпосередньо перед міграцією, багато птахів проявляють більшу активність, так званий «перелітний неспокій» (нім. Zugunruhe), та фізіологічні зміни, такі як накопичення жиру. На цю поведінку впливають не тільки зовнішні фактори. Виникнення перелітного неспокою навіть, у птахів, що виросли у неволі без будь-яких сигналів зовнішнього середовища, таких як скорочення світлового дня або зниження температури, вказує на роль генетично закодованих річних ритмів, що контролюють міграцію. Також птахи, що виросли у неволі, проявляють переважний напрямок польоту, що відповідає природному напрямку міграції, інколи навіть, роблячи зміни напрямку польоту, що відповідають природним.
Орієнтація і навігація
Навігація під час перельоту заснована на різноманітних органах відчуття. Багато птахів використовують Сонце в якості компаса. Використання Сонця для вибору напрямку польоту вимагає здатності до компенсації залежно від часу дня. Крім того, навігація може засновуватися на здатності відчувати магнітні поля або використовувати візуальну інформацію.
Більшість міграційних птахів зазвичай дещо розосереджуються ще молодими птахами, не повертаючись після зимування точно до місця народження. Проте дещо пізніше вони формують прив'язку до певних потенційних місць гніздування і зимування. Як тільки така прив'язка здійснюється, птах починає відвідувати саме ці місця щороку.
Здатність птахів до навігації протягом міграцій не може бути пояснена виключно генетичною програмою, навіть, за допомогою використання факторів навколишнього середовища. Здатність успішно здійснювати довгі міграції ймовірно може бути пояснена лише з врахуванням когнітивних можливостей птахів та можливості розпізнавання місць завдяки пам'яті. Супутникове слідкування за міграціями денних хижих птахів, таких як скопи (Pandion haliaetus) і осоїди (Pernis), показало, що старіші особини під час польоту краще роблять корекції на вітер.
Як показує наявність річних циклів, існує значна генетична складова у виборі часу та маршруту перельоту, проте ця програма може змінюватися під дією зовнішніх факторів. Цікавим випадком такої зміни, що виникає в результаті географічного бар'єра, є зміна міграційного маршруту деяких центральноєвропейських чорноголових кропив'янок (Sylvia atricapilla), що мігрують на захід і зимують у Великій Британії замість перельоту через Альпи.
Міграційні птахи також можуть використовувати електромагнітні механізми для орієнтації, зараз запропоновані два таких механізми: один природжений, а інший засновується на власному досвіді. Молодий птах протягом своєї першої міграції летить у вірному напрямку, зважаючи на магнітне поле Землі, проте не знає тривалості польоту та розташування природних бар'єрів. Вважається, що ця магнітна чутливість виникає за допомогою механізму радикальних пар (англ. radical-pair mechanism), при якому хімічні реакції в певних пігментах, чутливих до червоного та інфрачервоного світла, змінюються під дією магнітного поля. Хоча цей механізм і працює виключно протягом світлого часу доби, він не використовує положення Сонця. Молоді птахи, що використовують лише цей механізм, подібні бойскаутам з компасом, але без карти, поки вони не звикнуть до маршруту і зможуть використовувати інші методи орієнтації. З досвідом вони вивчають різноманітні ознаки ландшафту та зв'язують ці ознаки із силою та напрямком магнітного поля, вважається, що таке зв'язування відбувається за допомогою використання кристалів магнетиту в тригемінальній системі, яка повідомляє птаха про силу магнітного поля. Протягом подорожі між північними та південними регіонами сила магнітного поля змінюється залежно від широти, що дозволяє птаху визначити, коли він вже досяг місця призначення. Недавні дослідження також встановили зв'язок між оком птаха і «кластером N», частиною переднього мозку, активної протягом орієнтації, у результаті чого висувається думка, що птахи можуть «бачити» магнітне поле Землі.
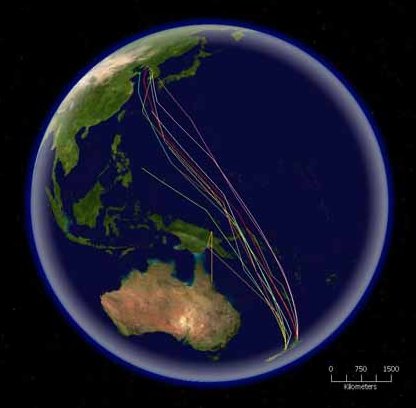{kind=link}
Маршрути малих грициків, мічених та простежених за допомогою супутника, на шляху на північ з Нової Зеландії довжиною біля 10 тис. км.
Помилки при міграції
Міграційні птахи можуть втратити шлях і опинитися поза межами свого звичайного ареалу. Найчастіше це трапляється в результаті перельоту далі місця призначення, часто на тисячі кілометрів, наприклад, коли птах опиняється на північ від місця гніздування. У результаті птах починає шукати шлях назад, здійснюючи так звану «зворотну міграцію», при якій генетична програма молодих птахів нездатна вірно працювати. Певні райони, через своє розташування, стали відомими як місця спостереження мігруючих птахів. Прикладами є Національний парк Пойнт-Пелі в Канаді і Спурн в Англії. Дрейфова міграція птахів, що збилися з курсу через вітер, приводить до «падіння» великої кількості мігруючих птахів у деяких прибережних місцях.
Штучне управління міграцією
У деяких випадках можливо штучно навчити зграю птахів міграційному маршруту, що необхідно при проведенні реінтродукції. Після експериментів з канадськими казарками (Branta canadensis), безпечному маршруту міграції вдалося навчити американських журавлів (Grus americana) за допомогою ультралегкого літака.
Еволюція та екологія виникнення міграції
Чи мігрує певний вид, залежить від ряду факторів. Найважливішим є клімат області, де птах гніздиться. Дуже небагато видів здатні прожити в жорстких умовах зими внутрішньої частини Канади або північної Євразії. Тому, наприклад, чорний дрізд (Turdus merula) міграційний у Скандинавії, але не є таким у м'якішому кліматі південної Європи. Джерело харчування також важливе. Більшість комахоїдних видів за межами тропіків мігрують на великі відстані та не мають вибору, крім того, щоб залишати гніздовий ареал взимку.
Часто різні фактори точно збалансовані. Європейська лучна трав'янка (Saxicola rubetra) та азіатська сибірська трав'янка (Saxicola maura) мігрують на великі відстані до тропіків, тоді як їхній близький родич європейська трав'янка (Saxicola rubicola) є осілим птахом на більшій частині свого ареалу і лише кочує на невеликі відстані з холодної півночі та сходу Європи. Перевагою осілих видів є додаткова можливість для розмноження.
Недавні дослідження пропонують, що горобцеподібні птахи, що мігрують на великі відстані, походять з південної та центральної Африки, а не північної півкулі. Вони по суті є південними видами, що мігрують на північ для гніздування, а не північними, що мігрують для зимування.
Також теоретичний аналіз показує, що обхідні маршрути, які збільшують довжину міграції до 20%, часто виникають у результаті адаптації, що птаху легше долати бар'єри з меншими жировими запасами. Проте, деякі види здійснюють міграції далекими від оптимальних обхідними маршрутами, що виникли в результаті історичного розповсюдження популяції. Наприклад, континентальні популяції дрозда Свенсона (Catharus ustulatus) летять далеко на схід через всю Північну Америку, повертаючи біля океану і досягаючи Південної Америки через Флориду. Вважається, що цей маршрут виник у результаті розширення ареалу зі східного узбережжя, яке відбулося біля 10 тис. років тому.
В інших випадках обхідні маршрути можуть бути викликані та надовго занесені у пам'ять через типові напрямки вітру, наявність хижаків або інші фактори.
Зміни клімату також впливають на час міграції, гніздування та інших подій життєвого циклу птахів, подібний же ефект має і зменшення численності популяцій.
{kind=link}
{kind=link}
Екологічні наслідки перельотів птахів
Міграція птахів також допомагає пересуванню інших видів, наприклад ектопаразитів, таких як кліщі (Acarina) і воші (Phthiraptera), які у свою чергу можуть переносити мікроорганізми, у тому числі збудників людських захворювань. Значну увагу міграційні птахи привернули у зв'язку з розповсюдженням пташиного грипу в 2006–2007 роках, проте з'ясувалося, що міграційні птахи не становлять особливої загрози, тоді як імпорт домашніх птахів має набагато більший ефект. Деякі віруси, проте, дійсно можуть переноситися птахами без помітного ефекту на здоров'я самого птаха, наприклад західнонільський вірус (WNV). Також міграційні птахи можуть грати роль у розповсюдженні насіння або спор рослин і планктону.
З початку досліджень міграції птахів з цією метою було розроблено велику кількість методів. Інколи методи, розроблені для зовсім інших пов'язаних процесів, стають неоцінимими для дослідження явища міграції.
Пряме спостереження
Найстарішим, найпростішим і найпоширенішим зараз методом дослідження міграції птахів є пряме спостереження. Розмір, колір, звуки і особливості польоту різних видів дають можливість як любителям, так і фахівцям робити висновки щодо їхньої міграції. Багато державних служб різних країн регулярно публікують результати таких спостережень. У сукупності, пряме спостереження надало більшу частину наших знань щодо міграції, але цей метод у значній мірі обмежується спостереженнями протягом світлої частини доби та наземними видами птахів.
«Місячне спостереження» — нічна модифікація методу прямого спостереження, що дозволяє спостерігати за видами, що мігрують уночі. До розробки методу в середині 20 століття даних про нічну міграцію майже не існувало. Цінна інформація може бути отримана за допомогою спостереження прольоту птахів на фоні повного Місяця за допомогою малосильних телескопів, що дають можливість підрахунку числа птахів, що перетинають його, так і напрямку їхнього польоту. Проте, оскільки фактичний відсоток неба, що спостерігається таким чином, невеликий (приблизно одна стотисячна площі неба), обсяг даних, що отримуються в результаті, також відносно незначний. Зазвичай під час сезону міграції вдається підраховувати біля 30 птахів за годину. Але той факт, що спостерігається навіть така кількість, свідчить про велике число птахів, що мігрують уночі.
Слуховий метод
Інший нічний метод спостереження, який дуже допомагає при опізнанні різних видів під час міграції, — використання параболічного відбивача з прикладеним мікрофоном для підсилення голосових сигналів птахів та приладом для їхнього запису. Цей пристрій може записувати звуки нічних міграційних птахів на відстані до 4 км в умовах безмісячної ночі, коли неможливе ніяке оптичне спостереження. Проте, незручністю методу є те, що при його використанні важко одразу визначити, що птах мігрує. До того ж існують деякі труднощі з упізнавання голосових сигналів через те, що голосові сигнали під час нічного перельоту можуть відрізнятися від сигналів, що їх подають птахи вдень. Крім того, птах може не подавати ніяких сигналів під час безпосереднього прольоту над зоною спостереження.
Збережені зразки птахів
Додатковий матеріал отримується за допомогою дослідження збережених залишків птахів разом з даними про час та місце їхнього збору. При використанні цього методу також важливо збирати певну кількість птахів у місцях їх гніздування та зимування, що дозволяє ототожнити окремі популяції одного виду птахів. Ці зразки порівнюють зі зразками, зібраними протягом міграції, пов'язуючи їх разом, що забезпечує впізнавання особин відомих популяції, незалежно від місця збору зразків. Хоча для збору зразків можуть використовуються птахи, вбиті мисливцями, важливим джерелом птахів, отриманих без їхнього зумисного знищення, є птахи, які зіштовхнулися з високими штучними спорудами або впали в результаті штормів та інших нещасних випадків.
Мічення
Поширеним способом є відловлювання птахів, яких потім мітять і випускають на волю неушкодженими. Спостереження отримані завдяки цим міткам надають багато корисних відомостей про пташині міграції. Було розроблено багато різних методів такого мічення, що дозволяють ідентифікувати окремих птахів. Найстарішим методом мічення є кільцювання птахів, при якому мітка прикріпляється до ніг птахів, їхньої шиї, крил або інших частин тіла. Щороку професійні біологи і добровольці, які працюють разом з ними, прикріпляють мітки до тисяч птахів, як міграційних, так і осілих. Кожна мітка містить порядковий номер і адрес наукової групи, до якої мітка повинна бути надіслана у випадку знаходження. Дані про кожного птаха та час мічення записуються, що дозволяє пізніше встановити факт його переміщення. Отримання великої кількості подібних даних дозволяє встановити багато деталей міграції помічених птахів.
Дані, отримані за допомогою кільцювання, надають таку інформацію, як час відправлення і прибуття до місця призначення, довжину пауз на міграційних шляхах для годування і відпочинку, співвідношення між погодними умовами і часом початку міграції, швидкості польоту індивідуальних птахів і ступінь регулярності, з якою індивідуальні птахи повертаються до характерних літніх або зимових районів, де вони мешкали в попередні роки. Крім того, ці дослідження дозволяють отримати дані щодо життєзабезпечення специфічних популяцій або їхньої уразливості до полювання.
Замість кільцювання інколи використовуються мічення за допомогою кольорової фарби або мічення за допомогою стабільних ізотопів водню або стронцію.
Радіоспостереження
Радіоспостереження або телеметрія — метод, що використовує невеликий радіопередавач, який видає періодичні сигнали з тіла мігруючого птаха. Радіоприймач, що може знаходитися на будь-якому транспортному засобі, наприклад на літаку або на штучному супутнику Землі, дозволяє простежити за цими радіосигналами і відстежити місцезнаходження мігруючого птаха. Одним з перших відомих прикладів використання цього методу було спостереження сірощокого дрозда (Catharus minimus) в роботі 1965 року. До птаха був прикріплений передавач вагою 2,5 г та його політ був простежений протягом понад 8 годин під час перельоту від Урбани (штат Іллінойс) до півночі озера Мічиган (що знаходиться на відстані 700 км від Урбани). Птах показав швидкість біля 80 км/год при вітрі 40 км/год, що допомагав птахові. Обмеженням методу радіотелеметрії, звичайно, є розмір передавача, який не має заважати польоту, і забезпечення транспортного засобу, що повинен триматися достатньо близько від птаха для відстеження сигналів. З початку радіотелеметричних досліджень спостерігається значний прогрес у технології, що забезпечив можливість спостерігати політ птахів за допомогою супутників. Проте, метод все ще використовується обмежено, оскільки декілька досліджень продемонстрували, що передавачі значно знижують шанси птаха на виживання.
Спостереження за допомогою радарів
Радари були розроблені для ідентифікації літаків та інших великих апаратів протягом другої світової війни, проте цей метод швидко став корисним і для досліджень, не пов'язаних з авіацією, зокрема досліджень перельотів птахів. Цей метод дозволяє спостерігати птахів незалежно від погоди та протягом будь-якого часу доби. Загалом метод можна розділити на кілька різновидів:
- загальний радар спостереження, подібний до радарів спостереження за літаками в аеропортах, що проглядає велику область і указує загальний час і напрям польоту великих об'єктів, зокрема зграй птахів;
- радарний запис польоту за допомогою «захоплення» об'єкта;
- доплерівський радар, що дозволяє вимірювати швидкість літаків і мас повітря в метеорології.
Використання радара у дослідженні міграції було неоцінимим у визначенні напряму і швидкості руху великих зграй птахів, дати й часу підправлення їх у подорож і загальної кількості птахів, особливо вночі. За допомогою методу було, зокрема, відкрито відносно значні рухи співочих птахів та інших маленьких сухопутних птахів, що мігрують над океанами, а не уздовж берегових ліній, і в напрямках, про які наземні спостерігачі і не підозрювали.
Дослідження орієнтації
Дослідження поведінки орієнтації традиційно здійснюються за допомогою різноманітних варіантів установки, відомої як воронка Ельмена, яка складається з круглої клітки з верхом, покритим прозорим склом або дротяним екраном таким чином, що небо залишається видимим, альтернативно установка розміщується в планетарії або в іншій системі, що дозволяє птахові бачити небо. Поведінка орієнтації птаха усередині клітки вивчається кількісно, використовуючи розподіл міток, що птах залишає на стінах клітки.
Інший метод, що використовується для цієї цілі, використовує напрям, в якому птах зникає на горизонті.
{kind=link}
Воронка Ельмена
Загрози та збереження птахів
Людська діяльність завдає значної загрози мігруючим птахам. Велике значення мають місця зупинок між місцями гніздування та зимування, зникнення яких у результаті людської діяльності не дає птахам можливості харчування під час перельоту. Знищення заболочених територій у результаті використання їх для потреб сільського господарства залишається найважливішою причиною загибелі птахів під час міграції.
Полювання уздовж маршрутів міграції в деяких випадках завдає дуже великих збитків популяціям птахів. Так, популяції білого журавля, що гніздяться в Сибіру, а зимують в Індії, практично зникли в результаті полювання на них під час польоту над Афганістаном і Середньою Азією. Останнього разу ці птахи спостерігалися у своєму улюбленому місці зимування, Національному парку Кеоладео, в 2002 році.
Високі споруди, такі як лінії електропередачі, вітряки, вітряні електростанції і прибережні нафтові платформи є частою причиною зіткнення з ними і загибелі міграційних птахів. Особливу загрозу мають освітлені вночі споруди, такі як маяки, хмарочоси, великі пам'ятники і телевізійні вежі, з вогнями, що мають запобігати зіткненню з ними літаків. Світло часто привертає птахів, що здійснюють міграцію вночі, подібно до того, як воно привертає нічних комах.
Концентрація птахів протягом міграції є причиною додаткової загрози для певних видів. Деякі найвидовищніші міграційні птахи вже вимерли, найвідомішим є мандрівний голуб (Ectopistes migratorius), зграї якого становили до 2 км завширшки і до 500 км завдовжки, пролітали кілька днів над однією ділянкою і налічували до мільярда птахів.
Охорона міграційних птахів ускладнена через те, що міграційні маршрути перетинають кордони різних країн і таким чином вимагають міжнародної співпраці. Для охорони перелітних птахів були укладені кілька міжнародних договорів, включаючи Договір про міграційних птахів 1918 року в Північній Америці (англ. Migratory Bird Treaty Act в США), Африкансько-Євразійський договір про охорону водоплавних птахів 1979 року (англ. African-Eurasion Waterbird agreement) і Боннська конвенція 1979 року (англ. Convention on Migratory Species).
Міжнародний день мігруючих птахів
Всесвітній день мігруючих птахів був започаткований у 2006 році з метою популяризації ідей про необхідність охорони мігруючих птахів та їх середовища існування. Цей день присвячено даті підписання 10 травня 1906 року Міжнародного Договору про охорону птахів. У 1927 році СРСР (в тому числі і Українська РСР) ратифікував цей договір. Всесвітній день мігруючих птахів організовується Секретаріатами Конвенції про збереження мігруючих видів диких тварин та Угоди про збереження афро-євразійських мігруючих водно-болотних птахів. Цей День відзначається щорічно у другу суботу та неділю травня.
У загальній схемі кругообігу речовин і енергії в біогеоценозах птахи посідають місце первинних, вторинних або навіть третинних консументів. Це означає, що вони споживають або первинну продукцію біогеоценозу у вигляді рослин, або живляться тваринами (рослиноїдними чи твариноїдними).
Політ птахів полегшує пошук корму. Птахи добувають його у найрізноманітніших місцях: на землі, гілках дерев, з квітів, на скелях, у воді, повітрі. У пошуках їжі вони можуть долати великі відстані. Наприклад, серпокрилець чорний добуває їжу за десятки кілометрів від гнізда. Шукаючи корм, він долає протягом дня до 1000 км.
За різноманітністю кормів птахів зазвичай розділяють на три групи.
- Поліфаги (всеїдні) живляться різноманітними рослинними і тваринними кормами. До цієї групи належить приблизно третина родин, у межах родини всеїдність більше притаманна тим видам, які більші за розміром. Наприклад, великі воронові птахи (крук, ворона тощо), великі мартини.
- Стенофаги — види що споживають однорідні корми та застосовують одноманітні способи добування їжі. Стенофагія спостерігається серед птахів порівняно рідко. До стенофагів можна віднести серпокрильців та ластівок, що живляться тільки літаючими комахами. Тільки рибою живляться пелікани, скопа та інші. Змієїд живиться майже виключно плазунами; африканський пальмовий гриф Gynohierax angolensis вживає в їжу переважно плоди декількох пальм; американський шуліка Rostrhamus sociabilis живиться лише одним видом молюсків. Нектар квітів обмеженої кількості рослин вживають деякі види колібрі, медососи, гавайські квіткарки (Drepanididae).
- Більшість птахів належить до проміжної групи, вони вживають досить широкий набір кормів. Так, багато горобцеподібних живляться як різноманітними комахами, так і насінням. Рибою та різноманітними безхребетними живляться баклани, норці (Podiceps) та інші птахи; зеленими частинами рослин, ягодами, насінням та різноманітними безхребетними — куроподібні, тукани (Ramphastidae), птахи-носороги.
За складом кормів птахів поділяють на декілька екологічних груп.
{kind=link}
Будова дзьоба птахів залежить від харчової спеціалізації
- Фітофаги — види, що живляться переважно рослинною їжею. Майже виключно листям живиться гоацин; гуси, лебеді, деякі качки, лиски живляться головним чином різноманітною прибережною та водною рослинністю. Переважно насінням живляться багато горобцеподібних — ткачикові, в'юркові, жайворонкові. Насінням, ягодами та плодами живляться багато голубів, папуг, туканів. Слід відзначити, що більшість фітофагів споживають переважно найкалорійніші частини рослин — насіння та плоди.
- Зоофаги живляться переважно тваринною їжею. Майже третина родин птахів виключно або переважно комахоїдні (ентомофаги). Багато водних та навколоводних видів живляться переважно рибою (іхтіофаги). Це гагари, веслоногі, великі чаплі, крехи, деякі рибалочки, скопа, деякі орлани тощо. Багато соколоподібних та сов належать до міофагів (живляться дрібними гризунами); яструби, соколи сапсан та підсоколик великий є переважно орнітофагами (живляться птахами). Невелика кількість птахів належить до малакофагів (живляться переважно молюсками) — це пухівка, кулик-сорока тощо. До герпетофагів (живляться земноводними та плазунами) належать птах-секретар, змієїд та деякі інші.
Однак такий поділ за складом кормів значною мірою є схематичним. Адже за наявності масових легко доступних кормів, ними починають живитись і ті види, які зазвичай їх не вживають. Так, під час нашестя сарани у степу нею живляться не лише ентомофаги, а також качки, кулики, мартини, круки та багато інших птахів. У птахів спостерігаються сезонні зміни у живленні. У багатьох птахів з великими ареалами чітко виражена географічна мінливість живлення, що визначається наявністю та доступністю кормів. Доволі різкі коливання спектру живлення обумовлені доступністю кормів у різні роки. Доволі чітко виявляється індивідуальна перевага у виборі кормів. Практично у всіх птахів деякою мірою помітна вікова зміна кормів.
Способи добування кормів у птахів не дуже різноманітні. Більшість видів бере здобич дзьобом. Відповідно до харчової спеціалізації форма та розміри дзьоба варіюють в широких межах. Не менш різноманітна і форма язика, який у багатьох видів допомагає не тільки у заковтуванні їжі, але також бере участь у її захопленні та утримуванні. Так, у дятлів язик дуже висувається та має гострі шипи на кінці, якими птах намацує личинку та витягує її. Соколоподібні та сови ловлять здобич переважно лапами. Для птахів описано також використання певних «інструментів». Галапагоський в'юрок Camarhynchus pullidus, тримаючи у дзьобі голку кактуса, колупає нею у тріщинах кори, виганяючи комах назовні, а потім хапає їх дзьобом.
Високий рівень обміну речовин у птахів пов'язаний з переробкою великої кількості їжі. Великі тепловитрати у дрібних видів призводять до того, що вони потребують відносно більше їжі. Так, маса їжі, що споживається дрібними горобцеподібними за добу, становить 50—80% їхньої маси. Великий яструб масою близько 1,5 кг потребує лише 150–200 г м'яса, проте спіймавши велику здобич може з'їсти до 800–1000 г. Здатність до голодування залежить від розмірів птаха та його фізіологічного стану. Дрібні птахи гинуть без їжі вже через 15—30 годин, голуби — через 7—9 днів, великі орли та сови можуть голодувати до місяця
Значення в природі та житті людини
Взаємовідносини з людиною
Стосунки між людиною і птахами мають давню історію та є різноманітними за своєю природою. У більшості випадків людина використовує птахів для отримання м'яса, пір'я та пуху, а також в релігійних обрядах, для розваг та з метою комунікації (поштові голуби). Інколи між людиною і птахами спостерігаються мутуалістичні взаємовідносини, наприклад кооперація у пошуку меду між медоуказчиками (Indicatoridae) та африканськими народами, наприклад, борана. В інших випадках стосунки між людиною та птахами можна розглядати як коменсалізм, наприклад, стосовно хатнього горобця, який залежить від людської діяльності. Деякі види птахів завдають значної шкоди сільському господарству та становлять загрозу для літаків. Окремі види (свійський голуб) своїм послідом завдають шкоду пам'ятникам та архітектурним спорудам. У той же час діяльність людини (відстріл, забруднення навколишнього середовища, хижацтво з боку одомашнених котів, собак тощо) у багатьох випадках є причиною загибелі для багатьох птахів.
Птахи можуть брати участь у перенесенні на великі відстані багатьох хвороб (пташиний грип, сальмонельоз, орнітоз, кампілобактеріоз, лямбліоз та інші). Деякі з них, так звані зоонози, можуть бути небезпечними для людини.
Птахи можуть бути важливим індикатором стану довкілля, для чого застосовують різноманітні методи досліджень (моніторинг чисельності, видового складу, успішності гніздування тощо).
{kind=link}
Вирощування курей на фермі
Економічне значення
Багато птахів є родоначальниками свійських птахів, від яких одержують м'ясо, яйця, пух, що призвело до розвитку цілої галузі — птахівництва. Наразі це найбільша галузь для отримання тваринного білка для людини — 2003 року у світі було вироблено 76 млн тон м'яса та 61 млн тон яєць. Найпоширенішими птахами у птахівництві є кури, гуси, качки, індички, мускусні качки, цесарки, фазани, перепела та інші. Серед них виведено багато різноманітних порід.
Багато видів птахів належать до мисливських, на яких здавна полюють з метою отримання м'яса, проте на сьогодні у більшості регіонів (крім слабко розвинених країн) полювання здебільшого є розвагою. Найважливішими мисливськими птахами в Європі та Північній Америці є водоплавні — різні види качок та гусей, а також кулики, голуби, фазани, перепілки. Деякі види нелітаючих птахів, на яких люди полювали з метою отримання м'яса, було винищено (дронт, безкрила гагарка та ін.) Збирання пташенят буревісників залишається популярним в Австралії та Новій Зеландії.
{kind=link}
Балабан — один із видів, що утримують для соколиного полювання
Іншим комерційно значимим продуктом, що отримують від птахів, є пір'я (особливо пух гусей та качок), яке використовується для виготовлення одягу. Гуано морських птахів використовується як добрива, що містить велику кількість фосфатів та сполук Нітрогену. Тихоокеанську війну інколи називають війною за гуано, оскільки однією з стратегічних цілей було отримання контролю над запасами гуано.
Багато видів птахів одомашнені з практичною метою та як хатні тварини. Яскраво забарвлені птахи (папуги, майни) утримуються в неволі, що нерідко призводить до нелегальної торгівлі окремими видами, які перебувають під загрозою вимирання. Соколи, орли та баклани тривалий час застосовувалися, відповідно, для полювання та риболовлі. Поштові голуби, принаймні з початку нашої ери до Другої світової війни, мали важливе значення для комунікації. Наразі згадане застосування птахів частіше розглядається як хобі та розваги, а також як спорт (змагання голубів).
Мільйони людей у світі захоплюються спостереженнями за птахами (англ. birdwatchering), що є однією з формою відпочинку та туризму. Багато людей приваблюють птахів до своїх домівок, вивішуючи годівниці взимку та різноманітні штучні гнізда в період гніздування.
{kind=link}
Спостереження за птахами у Великобританії
Роль у природних екосистемах
Птахи є важливими складовими різноманітних екосистем, входять до ланцюгів живлення та здійснюють безпосередній вплив на екосистеми. Особлива помітна роль птахів в екосистемах островів. Оскільки птахи заселяли острови набагато швидше ссавців; на таких островах птахи можуть повністю відігравати екологічну роль ссавців.
Деякі птахи, що живляться нектаром, є важливими запилювачами рослин. В окремих випадках птахи є єдиними запилювачами рослин. Тому у рослин та птахів-запилювачів нерідко спостерігається коеволюція. Багато видів, що живляться плодами, відіграють важливу роль у поширенні насіння. Насіння багатьох видів рослин проростає тільки у разі, якщо пройшло через травний тракт птахів.
Тривале гніздування морських птахів на одному місці приводить до концентрації посліду у вигляді гуано, що можуть збагачувати місцеві ґрунти.
Релігія, фольклор і культура
Птахи відіграють важливу роль у релігії, фольклорі та масовій культурі багатьох народів. У релігії птахи можуть відігравати роль посланників або священників, що веде до возведення їх у ранг богів. Наприклад, у скандинавській міфології пара круків Хугін та Мунін літають по всьому світу Мідгарду та сповіщають богу Одіну про те, що відбувається
Птахи були частиною культури і мистецтва з доісторичних часів, їх використовували у ранньому печерному живописі. Птахи є важливим образом у поезії, наприклад Гомер включає соловейка до своєї Одісеї.
Сприйняття різних птахів нерідко відрізняється в різних культурах. Сови часто асоціюються з невдачею або смертю в частині Африки, але вважаються символом мудрості на більшій частині Європи.
.jpg){kind=link}
«Три птахи», друкована графіка, 15-е ст. Німеччина
Охорона птахів
Понад 100 видів птахів зникло протягом історичного часу. Найдраматичнішим скорочення видового багатства було в результаті антропогенної діяльності, ці втрати оцінюються в 750–1800 видів, що спостерігалося протягом колонізації людиною островів Меланезії, Полінезії та Мікронезії. Протягом останніх десятиліть багато видів птахів скорочують свою чисельність. За оцінками міжнародної організації Birdlife International та МСОП 2009 року під загрозою зникнення перебувало 1227 видів (англ. threatened species), що становить понад 10% світової фауни птахів.
87 видів птахів включено до третього видання Червоної книги України (2009).
Основною причиною, яка становить загрозу для птахів, вважають втрату біотопів. Інші загрози включають надмірне полювання, загибель у риболовецьких знаряддях (сітках, знаряддях ярусного лову), забруднення навколишнього середовища, (включаючи забруднення нафтою та використання пестицидів), конкуренція та хижацтво з боку інтродукованих видів та зміни клімату.
Завдяки природоохоронним зусиллям урядів та громадських організацій вдалося вберегти 16 видів птахів, які були на межі вимирання в період між 1994 та 2004 роками, включаючи каліфорнійського кондора.
Заходи щодо охорони різних видів птахів є індивідуальними. Проте основними напрямками охорони птахів є: охорона місць існування, розведення в неволі, реінтродукція та переселення.
{kind=link}
Каліфорнійський кондор — вид, який було врятовано від вимирання. Внаслідок природоохоронних заходів чисельність збільшилася з 22 до понад 300 особин
Джерела:
https://uk.wikipedia.org/wiki/%D0%92%D0%BE%D0%BA%D0%B0%D0%BB%D1%96%D0%B7%D0%B0%D1%86%D1%96%D1%8F_%D0%BF%D1%82%D0%B0%D1%85%D1%96%D0%B2
https://uk.wikipedia.org/wiki/%D0%9F%D1%82%D0%B0%D1%85%D0%B8#.D0.93.D0.BE.D0.BB.D0.BE.D1.81.D0.BE.D0.B2.D0.B8.D0.B9_.D0.B0.D0.BF.D0.B0.D1.80.D0.B0.D1.82_.D1.82.D0.B0_.D0.B9.D0.BE.D0.B3.D0.BE_.D1.80.D0.BE.D0.BB.D1.8C
https://uk.wikipedia.org/wiki/%D0%9C%D0%B0%D1%85%D0%BE%D0%B2%D1%96_%D0%BF%D0%B5%D1%80%D0%B0
https://uk.wikipedia.org/wiki/%D0%9F%D1%82%D0%B0%D1%88%D0%B8%D0%BD%D0%B5_%D0%B3%D0%BD%D1%96%D0%B7%D0%B4%D0%BE
https://uk.wikipedia.org/wiki/%D0%AF%D0%B9%D1%86%D0%B5_%D0%BF%D1%82%D0%B0%D1%85%D1%96%D0%B2
https://uk.wikipedia.org/wiki/%D0%9E%D1%81%D1%96%D0%BB%D1%96_%D0%BF%D1%82%D0%B0%D1%85%D0%B8
https://uk.wikipedia.org/wiki/%D0%9C%D1%96%D0%B3%D1%80%D0%B0%D1%86%D1%96%D1%8F_%D0%BF%D1%82%D0%B0%D1%85%D1%96%D0%B2
https://uk.wikipedia.org/wiki/%D0%9F%D1%82%D0%B0%D1%85%D0%B8